Общая характеристика миоценовой флоры
Систематический состав рассмотренных выше комплексов и характер распространения их по территории СССР дают представление о флоре диатомей миоценового времени как типично пресноводной, свойственной в основном обширным озерным бассейнам эвтрофного типа. В миоцене такие озера были, по-видимому, широко распространены на территории Западно-Сибирской низменности, Прибайкалья, Забайкалья и юга Дальнего Востока.
В раннемиоценовое время флора диатомей отличалась довольно одно-образным родовым и видовым составом. В ней господствовали почти исключительно древние пресноводные виды рода Melosira (M. jouseana, M. ignota, M. praeislandica, M. praedistans, M. praegranulata). Развитие этих видов было столь обильным, что озерную диатомовую флору раннемиоценового времени можно назвать по преимуществу мелозировой с участием родов Tetracyclus, Fragilaria и Eunotia, а в Западной Сибири и рода Coscinodiscus. Большинство видов, бурно развивавшихся в раннемиоценовую эпоху, вымерло в основном в конце миоцена и раннем плиоцене, а некоторые характерные виды, такие как Melosira jouseana, M. ignota, Coscinodiscus lobatus, C. trapeciformis, по-видимому, исчезли уже в середине миоценового времени.
В позднемиоценовое время флора диатомей становится более разно-образной. В ней продолжают господствовать древние пресноводные виды Melosira, исключая вымерших M. jouseana и M. ignota, но заметную роль приобретают также M. italica var. italica et var. semilaevis, M. scabrosa, M. undulata. В озерах Западной Сибири и Прибайкалья наряду с ними обильно развиваются и древние пресноводные виды рода Coscinodiscus: C. gorbunovii, C. sibericus, C. miocenicus, которые на юге Дальнего Востока появляются лишь в плиоцене. Одновременно с указанными видами Melosira и Coscinodicus большое значение приобретают Tetracyclus, Fragilaria, Eunotia и Achnanthes. Они представлены своеобразным комплексом форм, часть которых полностью вымерла к концу миоцена (Fragilaria miocenica var. chankensis, Eunotia nikolskiae var. nikolskiae et var. gracilis, submonodon f. fossilis, Achnanthes miocenica), а часть сохранилась в плиоценовой флоре (круг форм Tetracyclus ellipticus, Fragilaria miocenicay Eunotia japonica, E. polyglyphoides и некоторые другие).
Таким образом, пресноводная озерная флора диатомовых водорослей СССР в миоценовое время в целом не отличалась большим систематическим разнообразием. Массовое развитие свойственно было лишь ограниченному числу родов и видов в основном из подкласса Centrophycidae (роды Melosira и Coscinodiscus). Эти виды в настоящее время полностью вымерли. Из пеннатных диатомей вполне очевидно доминирование представителей порядков Araphales, Raphidiales и Monoraphales, отличающихся более простым строением шва и панциря.
Для озерной флоры миоценового возраста в СССР установлено около 100 видов, разновидностей и форм диатомей, принадлежащих к 27 родам. Более половины характерных представителей этой флоры ныне вымерло. По-видимому, в дальнейшем число таксонов пресноводных миоценовых диатомей будет увеличиваться за счет изучения еще очень слабо исследованных районов европейской части СССР, Сибири, Северо-Востока, Тихоокеанской области.
Несмотря на своеобразие систематического состава комплексов диатомей миоцена в отдельных регионах и в целом по Советскому Союзу, они вполне сопоставимы между собой и с одновозрастными комплексами за пределами СССР. В каждом из комплексов присутствует значительное число эндемичных форм, свойственных древним водоемам только данного района, но наряду с этим на обширных территориях существовала определенная группа общих видов диатомей, дающая возможность для широких корреляций.
Результаты сравнительного изучения миоценовых пресноводных диатомей (табл. 16) показали, что дальневосточный комплекс диатомей ранне-среднемиоценового возраста по систематическому составу сравним пока только с комплексом раннего миоцена Забайкалья. Общими для них являются такие характерные формы, как Melosira jouseana, M. praedistans, M. praegranulata, M. praeislandica, Tetracyclus ellipticus var. ellipticus, Gomphocymbella miocenica, Didymosphenia geminata var. curvata и некоторые другие - всего 20 форм, что составляет значительный процент при относительно небольшом систематическом разнообразии комплексов диатомей раннего миоцена.
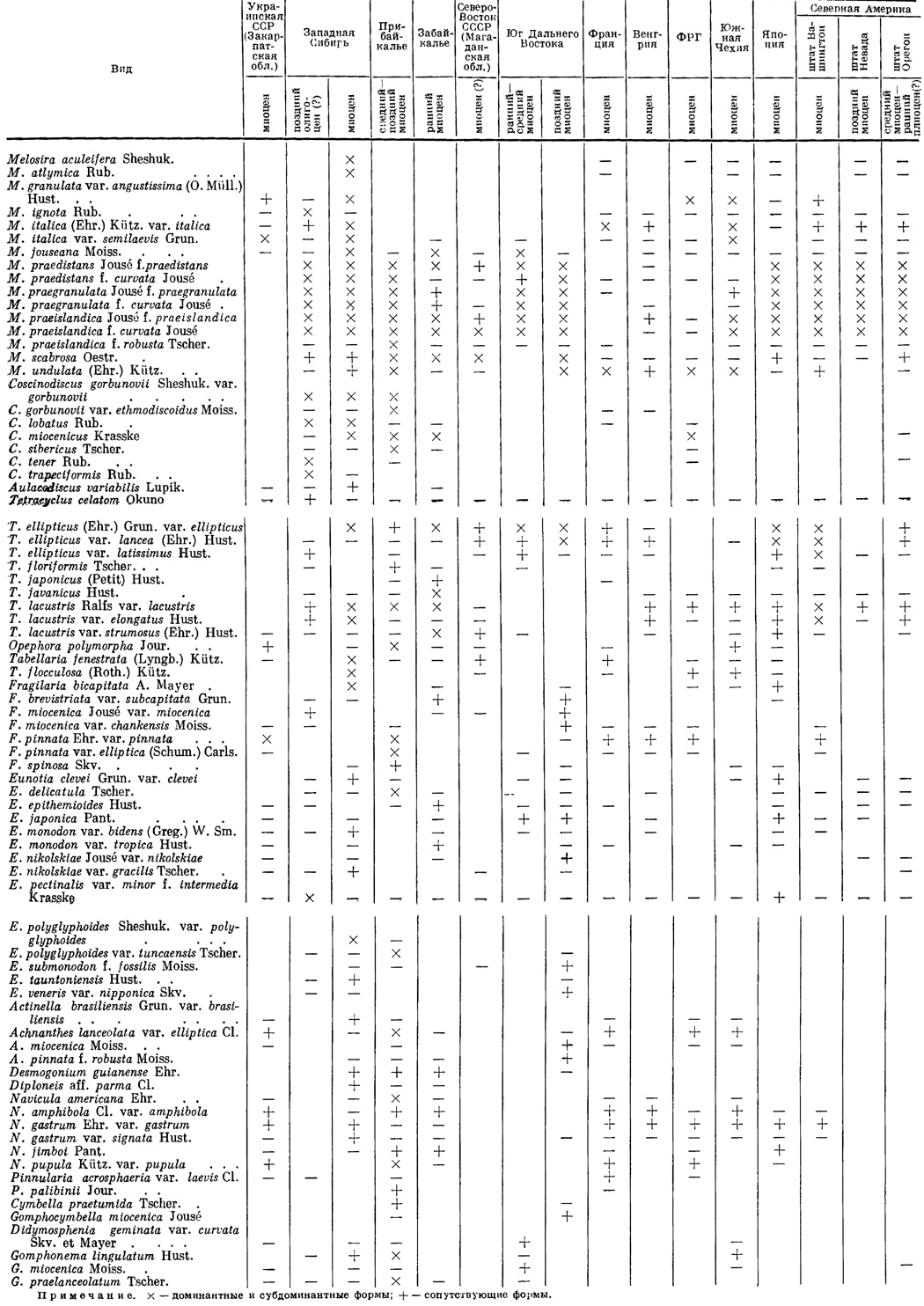
Таблица 16. Распространение пресноводных диатомовых водорослей, характерных для миоцена СССР
Эти комплексы имеют также некоторые родственные связи с комплексом диатомей из абросимовской свиты Западной Сибири. Сближает их доминирующее положение грубопанцирных видов рода Melosira и типичного для миоцена M. jouseana. Однако имеются между ними и существенные отличия. Основное из них: доминирование в абросимовском комплексе M. atlymica, Coscinodiscus lobatus, Aulacodiscus variabilis, которые отсутствуют как в дальневосточном, так и забайкальском комплексах.
Комплекс диатомей позднего миоцена юга Дальнего Востока обнаруживает ряд общих черт со средне-позднемиоценовым комплексом диатомей Прибайкалья. Доминируют в них грубопанцирные пресноводные виды рода Melosira. Высоких количественных оценок достигают также M. scabrosa, M. undulata var. undulata. Общими являются характерные формы Tetracyclus ellipticus var. ellipticus et var. lancea. Существенное различие между этими комплексами заключается в наличии в Прибайкалье целого ряда форм с высокой численностью, которые на Дальнем Востоке заметное количественное участие приобретают только в плиоценовое время (Coscinodiscus gorbunovii var. gorbunovii et var. ethmodiscoidus, Tetracyclus lacustris var. lacustris, Navicula americana, N. amphibola, Gomphonema lingulatum).
Сопоставление миоценовых комплексов диатомей СССР с одновозрастными пресноводными комплексами, известными за рубежом, свидетельствует о том, что наибольшее количество общих элементов имеют комплексы позднего миоцена юга Дальнего Востока и Прибайкалья и миоценовые комплексы Японии, о. Хоккайдо, в провинции Сэтана (Pantocsek, 1886-1892; Okuno, 1952а, 1958-1959) и п-ова Ното (Okuno, 1952а; Ichikawa a. oth., 1955). Здесь установлено 18 общих форм, среди них доминируют грубопанцирные виды рода Melosira (M. praedistans, M. praegranulata, M. praeislandica), описанные Окуно (Okuno, 1958-1959) как круг форм M. granulata. Почти повсеместно присутствуют также M. scabrosa, многочисленные формы Tetracyclus ellipticus, T. lacustris var. lacustris, Eunotia clevei var. clevei, E. japonica, E. praerupta. Одной из особенностей систематического состава миоценовой флоры Японии является присутствие в ней в значительном количестве планктонных видов Stephanodiscus niagarae var. minutulus и Coscinodiscus hokkaidoensis, неизвестных в миоцене Дальнего Востока и Прибайкалья. Кроме того, в миоценовых озерах Японии заметным развитием пользовались многочисленные виды Eunotia, Cymbella и Gomphonema, которые в СССР широко известны в пресных водоемах начиная с позднего плиоцена.
Родственные связи существуют также между комплексами диатомей миоцена юга Дальнего Востока и в несколько меньшей степени Прибайкалья и Западной Сибири с миоценовыми диатомовыми комплексами Северной Америки: штата Вашингтон (Van Landingham, 1964), Невада {Okuno, 1958) и Орегон (Moore, 1937). С миоценовой флорой штата Вашингтон установлено 17 общих форм, среди которых ведущее положение занимают группа грубопанцирных видов Melosira, Coscinodiscus miocenicus, круг форм Tetracyclus ellipticus и T. lacustris. Отличительной особенностью комплекса диатомей из штата Вашингтон является присутствие в его составе (до 60%) вида Melosira canadensis и в заметном количестве морского вида Coscinodiscus subtilis,* отмеченного К. Ломаном (см.: Moore. 1937) и для континентальных миоценовых отложений штата Орегон.
* (Достоверность определения Coscinodiscus subtilis требует еще тщательного подтверждения.)
Данные по позднемиоценовой озерной диатомовой флоре штата Невада весьма немногочисленны. Однако известно, что наиболее характерными элементами этой флоры опять-таки являются грубопанцирные виды Melosira, а также M. italica var. italica, Tetracyclus celatom, T. lacustris var. lacustris, преобладающие и в составе миоценовых комплексов пресноводных диатомей азиатской части СССР.
В комплексах диатомей среднемиоценового - раннеплиоценового (?) возраста штата Орегон в массе встречаются те же грубопанцирные виды Melosira, описанные здесь К. Ломаном (Moore, 1937) как круг форм M. granulata и M. solida. Эти виды в значительной мере образуют озерные диатомиты, широко распространенные в Северной Америке (Жузе, 1968а). Из других характерных форм общими с миоценовыми диатомеями СССР являются M. scabrosa, M. italica var. italica, круг форм Tetracyclus ellipticus и T. lacustris. Следует отметить, что К. Ломаном приводится не полный состав комплексов, а лишь доминантные формы и возраст их часто указывается лишь предположительно. В связи с этим и сопоставление комплексов возможно лишь в самых общих чертах.
Имеется некоторое сходство состава диатомей в миоценовых озерах Дальнего Востока, Прибайкалья и Южной Чехии (Rehakova, 1963, 1965). Общими для них являются Melosira praeislandica f. praeislandica et f. curvata, M. praegranulata f. praegranulata, M. italica var. italica, M. undulata var. undulata, Tetracyclus lacustris var. lacustris, составляющие группу доминантных видов. Однако широко распространенные в миоценовых озерах Чехии Opephora martyi var. martyi, Fragilaria construens var. construens, Navicula amphibola, N. bituminosa, N. gastrum, N. scutelloidesr Gomphonema lingulatum на Дальнем Востоке в заметном количестве от-мечены лишь в комплексах плиоценового времени.
Миоценовые комплексы пресноводных диатомей Западной Европы, известные во Франции (Heribaud, 1902-1908; Lauby, 1910; Ehrlichr 1966), Венгрии (Grunow, 1882; Pantocsek, 1903-1905; Palik, 1959; Hajos, 1959; Hajos, Palfalvy, 1963) и ФРГ (Krasske, 1934), по составу видов не сходны с одновозрастными комплексами СССР. Некоторое сходство имеется лишь среди видов широкого географического распространения, которые характеризуются длительным существованием (от миоцена доныне) и, как правило, не имеют прямого значения при установлении возраста комплексов. Кроме того, следует учесть, что миоценовые комплексы пресноводных диатомей европейской части СССР изучены еще мало и с западноевропейскими приходится сравнивать комплексы миоцена в основном азиатской части СССР, которые развивались в иной биогеографической обстановке.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'