Глава VIII. Диатомовые водоросли мезозоя (Стрельникова Н. И.)
Диатомеи юрского возраста достоверно не найдены. То, что до сих пор относилось к юрским диатомеям, в настоящее время подверглось пересмотру. По данным Дефляндра (Deflandre, 1959), юрские виды Ротплетца (Rothpletz, 1896) Pyxidicula bollensis, P. liassica - не диатомеи и относятся к Schizosphaerelidae (ископаемые известковые микрофоссилии неопределенного систематического положения), органическая оболочка которых в процессе фоссилизации минерализуется (кальцифицируется). Два вида диатомей: Compositus evgenii Veksch., Stephanopyxis parentesVeksch. - из позднеюрских отложений Западно-Сибирской низменности, описанные В. Н. Векшиной (1960), требуют дополнительных исследований и подтверждения. В верхнеюрских отложениях Подмосковного бассейна А. П. Жузе (Жузе, Шешукова-Порецкая, 1963 78) нашла остатки организмов, которые были условно отнесены ею к диатомеям. Однако ее дополнительные исследования показали их принадлежность к радиоляриям.
Отсутствие диатомей в морских юрских отложениях в первую очередь следует объяснять плохой сохраняемостью их панцирей. Не исключается возможность находок диатомей в отложениях юрского возраста, поиски их следует продолжать.
Сведения о диатомеях, существовавших в течение длительного отрезка времени, с юры до палеогена, далеко не полные; особенно мало данных о раннемеловых диатомеях.
Раннемеловые диатомеи. В альбских глинах по р. Выши в Пензенской обл. А. П. Жузе (1949а) обнаружила многочисленные сильно деформированные остатки диатомей, среди которых удалось определить Gladius sp., Stephanopyxis sp., Coscinodiscus sp. В альбских фосфоритах около г. Ганновера в Западной Европе Форти и Шульц (Forti, Schulz, 1932) нашли и описали 10 видов диатомей: Stephanopyxis mirabilis, S. turris var. campanella, Craspedodiscus incurvus, Dactyliosolen priscus, Ditylum cornutum, Gladius antiquus, Isthmia fortii, Ktenodiscus germanicus, Pyrgodiscus antiquus, Radiodiscus cretaceus. Это пока единственная находка хорошо сохранившихся диатомей в точно датированных отложениях альба. Все известные в настоящее время диатомеи раннемелового возраста относятся к подклассу Centrophycidae.
Позднемеловые диатомеи. Значительно больше известно о диатомеях позднемелового возраста, панцири которых обнаружены в отложениях туронского, кампанского, маастрихтского ярусов и в нерасчлененных отложениях сенона.
Остатки диатомей туронского возраста найдены в Чехии (Wiesner, 1936) и во Франции (Deflandre, 1941). Это полурастворившиеся, часто пиритизированные створки Stephanopyxis, Thalassiosira, Coscinodiscus, Triceratow. Описанная Мюллером Actinoclava frankei из турона Вестфалии (Miiller, 1911), по данным Дефляндр-Риго (Deflandre-Rigaud, 1962), является склеритом ископаемой голотурии и называется теперь Myriotrochites frankei Frizz, et Exl.
Хорошо сохранившиеся створки диатомей описаны Шульцем (Schulz, 1935) из поровых каналов губок сенонского возраста около г. Гдыни в Польше. Богатейшая флора диатомей из отложений верхнего мела из-вестна во многих местах Западной Сибири (Жузе, 1948, 1949а, 19496, 1951а, 1955а; Кротов, 1957а, 19576, 1959; Возженникова, 1960; Кротов, Шибкова, 1961; Шибкова, 1961; Векшина, 1961а, 19616, 1962; Стрельникова, 1964, 1965а, 19656, 1966а, 19666, 1966в, 1968; Глезер, 1966).
Диатомеи маастрихтского возраста найдены в диатомитах формации Морено в Калифорнии (Наппа, 1927а, 1934; Long a. oth., 1946; Barker, Meakin, 1946-1947). Кроме того, полурастворившиеся и пиритизированные ядра диатомей обнаружены Вейдеманом (Weidemann, 1964) в Швейцарии, в предгорьях Альп, в отложениях, предположительно датируемых Маастрихтом.
На территории СССР позднемеловые диатомеи встречены в нескольких районах (рис. 7, 1-10). Впервые позднемеловые диатомеи были найдены и описаны А. П. Жузе (1948, 1949а, 19496) в Западной Сибири из верхнемеловых отложений Свердловской обл. (в бассейне р. Б. Актай Ново-Лялинского р-на, у с. Курьи на левом берегу р. Пышмы и по р. Сосьвеу пос. Гари). В дальнейшем диатомеи позднемелового возраста были найдены во многих местах восточного склона Полярного, Приполярного и Северного Урала (Жузе, 1951а; Кротов, Шибкова, 1961). Многочисленные прекрасно сохранившиеся створки диатомей слагают здесь толщи диатомитов, которые неширокой полосой тянутся вдоль Полярного и Приполярного Урала, достигая местами 200 м мощности. Южнее г. Ивделя и на Северном Урале остатки диатомей в верхнемеловых отложениях встречаются редко. Самое южное местонахождение диатомей: пос. Семиозерное в Тургайской низменности. Диатомеи известны также на Гыданском п-ове по р. Танаме.
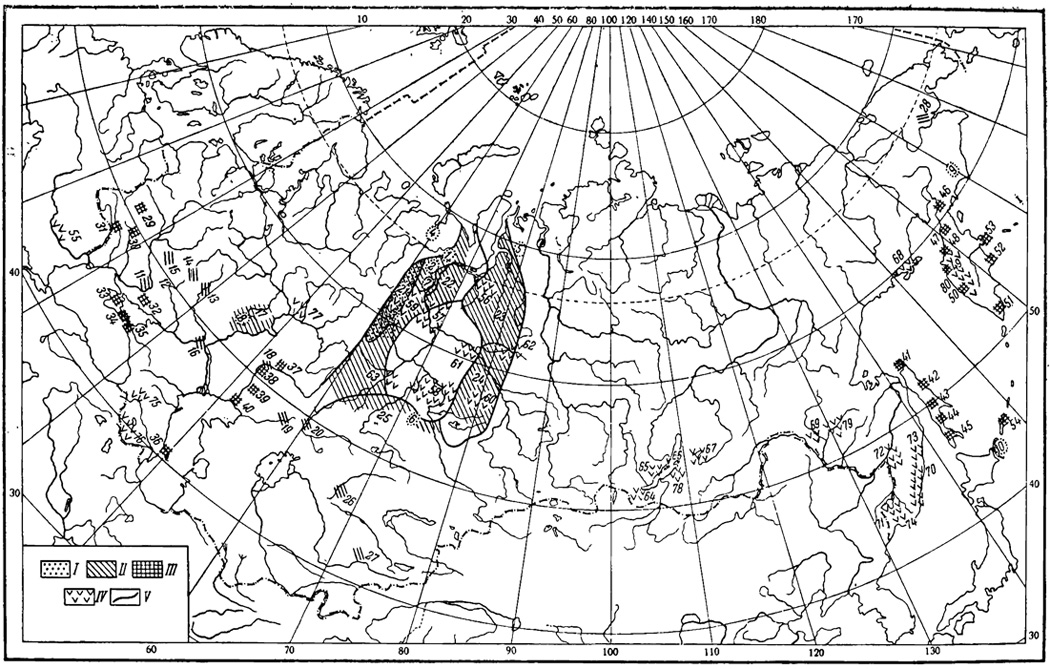
Рис. 7. Карта местонахождений позднемеловых (1-10), палеогеновых морских (11-28) и неогеновых (29-78) диатомей СССР. I - позднемеловые морские диатомеи: 1 - Мужинский Урал, 2 - восточный енлон Полярного Урала, 3 - восточный склон Приполярного и Северного Урала, 4 - западное Приуралье, г. Инта, 5 - Гыданский п-ов, р. Танама, 6 - пос. Семиозерное, Кустанайская обл., 7 - р. Сев. Сосьва, 8 - р. Мокина, 9 - р. Апука, Корякский нац. округ, 10 - о. Кунашир, Курильские о-ва. Л - палеогеновые морские диатомеи: 11 - Днепропетровская обл., 12 - Харьковская и Сумская обл., 13 - Воронежская обл., 14 - Курская обл., 15 - Черниговская обл., 16 - Волгоградская обл., 17 - Среднее Поволжье, 18 - окр. г. Уральска, 19 - басс. р. Эмбы, 20 - юго-запад Тургайской низменности, 21 - Приуральский фациальный р-н, 22 - Березово-Курганский фациальный р-н, 23 - Пур-Тазовский фациальный р-н, 24 - Кулундинско-Васюганский фациальный р-н, 25 - Приказахстанский фациальный р-н, 26 - Южный Казахстан, 27 - Ферганская долина, 28 - р. Анадырь. III - неогеновые морские исолоноватоводные диатомеи. Миоценовые диатомеи европейской части СССР: 29-30 - Хмельницкая обл., 31 - северо-запад Молдавской ССР, 32 - Запорожская обл., 33 - северо-восток Крымского п-ова, 34 - Керченский п-ов, 35 - Таманский п-ов, 36 - Апшеронский п-ов; плиоценовые диатомеи европейской части СССР: 37 - сел. Аксуат и Дарвинское, окр. г. Уральска, 38 - сел. Битик, Уральская обл., и район оз. Челкар, 39 - купол Матснкожа к северу от оз. Индер, Гурьевская обл., 40 - сел. Дарой-Мола, купола Теркобай и Бакланий, площадь Лиман и купол Камышитовый, Гурьевская обл.; неогеновые диатомеи Дальнего Востока. Сахалин: 41 - п-ов Шмидта, 42 - пос. Катангли, pp. Чамгу и Конги, восточное побережье Северного Сахалина, 43 - басс. р. Поронай, 44 - побережье зал. Терпения, Макаровский р-н, 45 - южная часть Южного Сахалина; Камчатка: 46 - Рекинникская губа, 47 - басс. р. Паланы, 48 - М. Непропуск, Тигильский р-н, 49 - утесы Энемтен, Тигильский р-н, 50 - Болынерецкая депрессия, 51 - басс. р. Опалы, 52 - Кроноцкий р-н, 53 - Усть-Камчатский р-н; Курильские о-ва: 54 - о. Итуруп. IV - неогеновые пресноводные диатомеи. Миоценовые диатомеи: 55 - Закарпатская обл., Виноградовский р-н, сел. Горобки, 56 - р. Правая Хета, басс. р. Надыма, 57 - басс. р. Назыма и правобережье р. Оби ниже устья р. Карымкары и у с. Подгорного, 58 - правобережье нижнего течения р. Иртыша, г. Тевриз Омской обл. и с. Ильинка Томской обл., бассейн нижнего течения р. Ишима, 59 - бассейны рек Сев. Сосьвы, Пелыма, Ирбита, Пышмы и Конды (pp. Супре и Б. Умытье), 60 - басс. р. Тары у с. Межевки, 61 - басс. р. Б. Югана, 62 - правобережье р. Тыма, урочище Кампасский Бор, 63 - окр. сел Новопетровки, Новой Троицы, Мизоново, Мичурино Тюменской обл., 64 - Прибайкалье, Тункинский р-н и Ахаликское буроугольное месторождение, 65 - оз. Нуха-Нур, Ангаро- Ленское междуречье, 66 - оз. Байкал, о. Ольхон, 67 - бассейн верхнего течения р. Витима, 68 - окр. г. Магадана, 69 - бассейн верхнего течения р. Зеи близ устья р. Томи, 70 - северная и северовосточная части хребта Сихотэ-Алинь, 71 - Южное Приморье, западное побережье оз. Ханка, 72 - Средне-Амурская депрессия; плиоценовые диатомеи: 64 - Прибайкалье, Тункинский р-н и Ахаликское буроугольное месторождение, 73 - район Советской Гавани, 74 - Южное Приморье, Приханкайская равнина, бассейны рек Суйфуна и Супутинки, 75 - Грузинская ССР, Ахалцихский р-н, у сел. Кисатиби, 76 - Армянская ССР, западное побережье оз. Севан и ущелье Ашри-дара у сел. Нурнус, 77 - Прикамье, бассейн нижнего течения р. Камы, 78 - юго-восточное побережье оз. Байкал, низовья р. Селенги, 79 - Амурская обл., басс. р. Арги, 80 - западное побережье п-ова Камчатка. V - границы фациальных районов
Во всех указанных выше пунктах встречена близкая по составу флора диатомей, что позволило уже первым исследователям (Жузе, 1948, 1949а, 19496; Кротов, 1957а, 19576; Кротов, Шибкова, 1961) выделить характерные для отложений верхнего мела виды: Melosira cretacea, Stephanopyxis schulzii, Coscinodiscus dissonus, Triceratium schulzii, Hemiaulus asymmetric cus, H. echinulatus, H. rossicus, Gladins speciosus. Среди диатомей доминирует Stephanopyxis schulzii. Возраст комплекса с S. schulzii вышеуказанные авторы принимали как сантон, сантон-кампан, кампан-маастрихт. В унифицированной региональной стратиграфической схеме верхнемеловых отложений Западно-Сибирской низменности (Решения. , 1969) породы, содержащие комплекс диатомей с S. schulzii, отнесены к верхней части березовского горизонта, который объединяет отложения коньяк- ского, сантонского и кампанского ярусов.
Детальное изучение диатомей из разрезов Полярного и Приполярного Урала, где верхнемеловые отложения содержат остатки руководящих групп моллюсков, дает основание датировать комплекс диатомей с S. schulzii кампанским временем (Стрельникова, 1965а, 1966в). Такая датировка принята по следующим соображениям: диатомиты залегают на породах с раннекампанскими аммонитами Scaphites cf. hippocrepis Decay, S. cuvieri Mort., Baculites acuminatum Glasun., B. cf. obtusus Glasun., что определяет нижнюю возрастную границу, и перекрываются в ряде разрезов известковистыми породами Маастрихта с Guadryiceras pigmaeum Glasun. В отложениях коньякского и сантонского ярусов диатомеи не найдены.
Флора диатомей кампанского века Западной Сибири представлена 33 родами и 145 видами и внутривидовыми таксонами. В составе флоры имеются виды обоих подклассов: Centrophycidae и Pennatophycidae. К этому времени сформировались существующие и ныне семейства порядков Discinales, Biddulphiales, Soleniales. Среди диатомей кампана наибольшим разнообразием в видовом отношении отличается сем. Biddulphiaceae из порядка Biddulphiales. К этому семейству относятся 5 родов с 54 видами и внутривидовыми таксонами. Из порядка Discinales известно 16 родов с 64 видами; из сем. Coscinodiscaceae - 8 родов с 38 видами; сем. Actinodiscaceae представлено 6 родами с 12 видами, a Eupodiscaceae - 4 родами с 14 видами. Порядок Soleniales представлен 1 видом. Из порядка Mediales найдено несколько видов, относящихся к семействам Anaulaceae, Rutilariaceae. Подкласс Pennatophycidae представлен 3 видами диатомей из порядка Araphales, сем. Fragilariaceae, однако их участие в составе флоры небольшое. Обильна группа спор неустановленного систематического положения; к этой группе отнесены 4 рода с 14 видами.
По видовому разнообразию и численности преобладают Stephanopyxis (12 видов), Triceratium (21) и Hemiaulus (20). Из рода Stephanopyxis наиболее характерными являются S. schulzii и близкий ему вид S. ornata. В 1 г диатомита насчитывается до 5 млн створок этих видов, что составляет около 10% от общего числа створок диатомей в породе (по подсчетам, в 1 г диатомита содержится 40-70 млн створок диатомей). S. schulzii свойствен лишь меловым отложениям, в более молодых породах палеогена он отсутствует. Есть лишь одно указание А. П. Жузе (1949а) о единичной находке створок S. schulzii в отложениях нижнесызранского яруса палеоцена Ульяновской обл., в диатомите у ст. Инза. Почти все остальные виды рода Stephanopyxis, за исключением S. antiqua, S. reticulata, S. uralensis, известны и в более молодых отложениях третичного возраста.
Можно сказать, что меловая флора - это царство Hemiaulus и Triceratium. Род Hemiaulus в палеогене представлен уже значительно меньшим числом видов, а Triceratium своего расцвета, по-видимому, достигает в палеоцене. Из рода Triceratium во флоре кампана преобладают S. indefinitum. В 1 г диатомита насчитывается до 5 млн створок этого вида. Для флоры кампана характерны также T. anissimovae, T. tesselum, T. uralense, T. synicum, хотя численность этих видов невелика. Что касается Hemiaulus, то 7 видов этого рода свойственны лишь меловой флоре, из них очень характерны H. asymmetricus, H. giganteus, H. sporalis, H. elegans. Последний продолжает существовать и в палеогене, но в мелу он имеет высокую численность (до 4 млн створок в 1 г диатомита) и характеризуется большой морфологической изменчивостью. Наибольшей численности в отложениях мела достигает H. rossicus (12 млн створок на 1 г породы) - вид, характерный также и для палеоцена.
Несколько меньшее значение по сравнению с Triceratium и Hemiaulus имеют Coscinodiscus (9 видов) и Melosira (4 вида). Из видов Coscinodiscus для меловой флоры характерен C. dissonus, не имеющий, однако, высокой численности. Из других видов единично встречаются C. mutabilis, C. si biricus, C. solidus, известные пока только в позднемеловой флоре. Наибольшей численности достигают C. symbolophorus (до 2 млн в 1 г) и C. wittianus (до 1 млн в 1 г). Первый вид имеет длительную историю существования, он известен и в современных морях.
Для меловой флоры очень характерен Gladius speciosus с формами: f. speciosus, f. aculeolatus, f. poratus. Это одна из наиболее крупных, часто встречающихся диатомей. Вид этот интересен тем, что для него удалось проследить процесс изменения морфологических признаков во времени и образование новых внутривидовых таксонов, переходных от f. speciosus через f. aculeolatus к f. poratus. Постоянно присутствуют в составе меловой флоры виды родов Pseudopodosira, Actinoptychus, Aulacodiscus, Biddulphia, Eunotogramma, но количественно они не обильны. Остальные роды представлены 1 или 2 видами, некоторые из них имеют высокую численность, например Goniothecium odontella. Среди диатомей есть несколько редких родов, виды которых встречаются единично: Actinodictyon, Monopsia, Glyphodiscus, Syndetocystis. Следует отметить наличие в меловой флоре видов из бесшовных пеннатных форм: Sceptroneis aff. caducea, S. dimorpha.
Позднемеловые диатомеи Западной Сибири были морскими. Выяснение экологических группировок позднемеловых диатомей представляет большие трудности и не может быть достаточно точным, так как большинство видов и многие роды вымерли, а точные морфологические критерии для отнесения того или иного не существующего ныне вида к группе неритических или океанических диатомей, бентосных или планктонных, недостаточны. Поэтому для вымерших родов и видов нельзя с уверенностью утверждать, был ли тот или иной вид бентосным или планктонным.
Преобладающие среди позднемеловых диатомей виды родов Hemiaulus, Triceratium, Stephanopyxis, по-видимому, были колониальными формами, так же как современные представители этих родов. В ископаемом состоянии встречаются обычно отдельные створки, редко можно наблюдать соединение 2-3 панцирей. Вероятно, большинство позднемеловых видов этих родов, судя по строению створок, имело колонии в виде цепочек. В составе меловых диатомей имеется ряд форм, которые представлены в виде спор неизвестных нам родов и видов. Таковы Pterotheca, Pseudopyxilla, Goniothecium, которые являются характерными элементами позднемеловой флоры диатомей. Некоторые из них имеют высокие оценки обилия. Наличие спор свидетельствует о том, что эти виды были планктонными и обитали в неритической области моря.
В течение кампанского века резких изменений в составе диатомей Западной Сибири не происходило. Массовые виды Stephanopyxis schulzii, Coscinodiscus dissonus, C. symbolophorus, Triceratium indefinitum, Hemiaulus asymmetricus, H. rossicus, H. elegans, Goniothecium odontella характерны для всей толщи отложений кампана (табл. XI). Однако небольшие изменения видового состава и численности диатомей на протяжении кампанского времени имели место. Различия состава диатомей отмечали А. П. Жузе (1951а), А. И. Кротов и К. Г. Шибкова (1961) и на этом основании выделяли несколько комплексов, но причина этих различий неясна. Жузе полагала, что эти комплексы разнофациальные, Кротов и Шибкова указывали на несколько разновозрастных комплексов. При послойном изучении 200-метровой толщи диатомитов на Приполярном Урале Н. И. Стрельниковой (1966в, 1968) удалось выделить три комплекса диатомей, последовательно сменяющих друг друга по разрезу.
Первый, наиболее древний комплекс приурочен к отложениям нижнего кампана. Характерными видами этого комплекса являются Melosira eretacea, M. ornata, Stephanopyxis antiqua, S. uralensis, Lepidodiscus elegans, Triceratium schulzii, Hemiaulus echinulatus, Pterotheca simplex (табл. XI). Особенно обильны Melosira cretacea, M. ornata, Hemiaulus echinulatus, Lepidodiscus elegans, которые в отложениях верхнего кампана встречаются единично и редко. Только в породах нижнего кампана встречены Stephanopyxis antiqua, Hemiaulus danicus, Pterotheca simplex, в вышележащих отложениях эти виды не обнаружены. Подобный комплекс диатомей прослежен на Полярном Урале (р. Сыня), на Мужинском Урале. Близкий по составу видов комплекс найден в районе Яро-Танамского междуречья на Гыданском п-ове (Стрельникова, 1966в).

Таблица XI. Комплексы кампапских (1-7) и раннекампанских (первый комплекс - 8-13) диатомей, Зап. Сибирь. 1 - Stephanopyxis schulzii Stein., 2 - S. ornata Schulz, 3 - Coscinodiscus dissonus Schulz, 4 - Hemiaulus asymrhctricus Jouse, 5 - Triceratium indefinitum (Jouse) Streln., 6 - Hemiaulus rossicus Pant., 7 - H. elegans (Heib.) Grun., 8 - Pterotheca simplex Streln., 9 - Melosira cretacea Jouse, 10 - Stephanopyxis antiqua Jouse, 11 - S. uraleruis Streln., 12 - Hemiaulus echinulatus Jouse, 13 - Triceratium schulzii Jouse
Второй (средний) и третий (молодой) комплексы диатомей приурочены к различным стратиграфическим горизонтам верхнего кампана. Второй комплекс характерен для диатомитов, залегающих в основании верхнего кампана. Он выделяется по появлению Stephanopyxis turris var. turris. В составе этого комплекса преобладают Melosira sulcata var. sulcata, Pseudopodosira simpla, Sceletonema polychaetum, Stephanopyxis turris var. turris, S. broschii, Triceratium anissimovae, Hemiaulus antiquus, H. kittonii, Gladius speciosus f. speciosus, Pterotheca aculeifera, P. evermannii (табл. XII). Все эти виды очень обильны в отложениях. Второй (средний) комплекс прослежен в отложениях верхнего кампана на Приполярном и Мужинском Урале.
Диатомиты, залегающие в основании верхнего кампана, вверх по разрезу постепенно переходят в диатомовые глины. Параллельно изменению литологии изменяется и состав видов диатомей. Из диатомовых глин, залегающих в верхах кампана, выделен третий (молодой) комплекс диатомей, характеризующийся следующими видами: Melosira sulcata var. sulcata, Pseudopodosira aspera, Stephanopyxis turris var. turris, S. lavrenkoi, S. reticulata, Aulacodiscus jouseae, Triceratium planum, Г. coronatum, Hemiaulus undulatus, Gladius speciosus f. aculeolatus, G. speciosus f. poratus, Pseudopyxilla rossica (табл. XII). Этот комплекс хорошо выделяется в разрезах по появлению Gladius speciosus f. aculeolatus и G. speciosus f. poratus. Третий (молодой) комплекс диатомей прослежен на Приполярном Урале и далее к югу на Северном Урале.

Таблица XII. Комплексы позднекампанских диатомей (второй комплекс - 1-7, третий комплекс - 8-13б), Зап. Сибирь. 1 - Melosira sulcata (Ehr.) Kütz. var. sulcata, 2 - Stephanopyxis turris (Grev. et Am.) Ralfs, 3 - Hemiaulus antiquus Jousé, 4 - Pterotheca evermannii Hanna, 5 - P. aculeifera Grun., 6 - Triceratium anissimovae (Jouse) Streln., 7a, 7б - Gladius speciosus Schulz f. speciosus, 8 - Pseudopodosira aspera Jousé, 9 - Stephanopyxis reticulata Long, Fuge et Smith, 10 - Aulacodiscus jouseae Streln., 11 - Triceratium planum Streln., 12 - Hemiaulus undulatus Jousé, 13a, 13б - Gladius speciosus Schulz f. poratus Streln
Находки диатомей в отложениях верхнего мела, за исключением Западной Сибири, крайне редки. Диатомеи, близкие по составу видов диатомеям восточного склона Урала, встречены в районе г. Инты (Западное Приуралье). В алевролитах, залегающих на песчаниках пермского возраста, обнаружены створки диатомей, среди которых преобладают Melosira ornata, M. sulcata var. sulcata, Stephanopyxis schulzii, S. reticulata, S. turris var. turris, Coscinodiscus dissonus, Triceratium indefinitum, T. excavatum, Hemiaulus rossicus, Goniothecium odontellia, Gladius speciosus f. poratus. Большое количество створок Gladius speciosus f. poratus позволяет сблизить этот комплекс диатомей с третьим комплексом восточного склона Урала.
В Поволжье, по р. Мокше в Пензенской обл., в зеленовато-сером песке найдены разнообразные диатомеи хорошей сохранности, по составу видов близкие западносибирским комплексам кампана. Здесь преобладают Stephanopyxis schulzii, Coscinodiscus dissonus, C. wittianus, C. symbolophorus, Melosira cretacea, M. ornata, Lepidodiscus elegans, Syndetocystis sp., noptychus heterostrophus, Triceratium schulzii, Hemialus echinulatus, if. rossicus, Gladius speciosus и др. Небольшие отличия систематического состава не позволяют пока уверенно датировать эту флору кампанским возрастом, хотя различия эти могут быть и не возрастными, а фациальными. Около г. Кадышево (Ульяновская обл.), из окремненного мергеля сантон-кампанского возраста выделены панцири Coscinodiscus sp., Gladius sp.
А. П. Жузе (Жузе, Шешукова-Порецкая, 1963) указывает на наличие диатомей в верхнемеловых отложениях о. Кунашир (Курильские о-ва). Панцири диатомей плохой сохранности найдены в кремнисто-известковистом алевролите маастрихтского возраста в бассейне р. Апуки (Корякский нац. округ). Состав диатомей из этих отложений отличается от комплекса кампана Западной Сибири; имеются общие виды с диатомеями формации Морено в Калифорнии. Среди диатомей в бассейне р. Апуки удалось определить такие характерные для Маастрихта Калифорнии виды, как Coscinodiscus morenoensis, Trinacria deciusi.
Из 55 видов диатомей, найденных Шульцем в спонголитах сенонского возраста около г. Гдыни на берегу Гданьской бухты (Schulz, 1935), 35 яв-ляются общими с кампанскими диатомеями Западной Сибири. Массовыми формами в обоих комплексах являются Stephanopyxis schulzii, Hemiaulus rossicus. Для обоих комплексов характерны Melosira sulcata, Stephanopyxis turris, Coscinodiscus dissonus, Arachnoidiscus ehrenbergii, Actinoptychus heterostrophus, A. simbirskianus, Aulacodiscus archangelskianus, A. lahusenii, A. septus, Triceratium schulzii, Hemiaulus elegans, Gladius speciosus f. speciosus, Pterotheca aculeifera и др. Сходство систематического состава диатомей кампана Западной Сибири с сенонскими диатомеями Гданьской бухты А. П. Жузе (1949а) объясняет существованием сообщения между западносибирским и среднеевропейскими морями в течение сенона.
Диатомеи Маастрихта Калифорнии значительно отличаются от кампанской флоры Западной Сибири. По данным Хэнна (Наппа, 1927а, 1934) и Лонга, Фьюдже, Смита (Long a. oth., 1946), диатомеи формации Морено около г. Коалинга в Калифорнии представлены 130 видами, разновидностями и формами, относящимися к 45 родам. В маастрихтском комплексе диатомей Калифорнии в отличие от кампанского Западной Сибири количественно преобладают роды порядка Discinales, среди которых по разнообразию видов доминируют Coscinodiscus (19 видов), Aulacodiscus (16), Triceratium (11), Actinoptychus (10), Auliscus (10), Biddulphia (10), Trinacria (8), Stephanopyxis (4), Hemiaulus (1). Но 18 видов являются общими с кампанскими диатомеями Западной Сибири, например Stephanopyxis barbadensis, S. reticulata, S. turris, Goniothecium odontella, Pterotheca evermannii и единично Actinodictyon weissflogii, Aulacodiscus archangelskianus, A. striaius, Monopsia mammosa, Eunotogramma bivittatum. Различия в систематическом составе диатомей Западной Сибири и Калифорнии, по-видимому, объясняются не только разным возрастом, большое значение имели различия физико-географических условий местообитания.
При сопоставлении диатомей позднего мела Советского Союза с одно- возрастными диатомеями, известными за рубежом, намечаются две группы. Эти группы диатомей территориально совпадают с двумя палеобиогеографическими областями, выделенными при районировании по брахиоподам,. аммонитам, белемнитам и другим группам организмов (Макридин, Кацг 1966). Западносибирские, интинские и пензенские, а также гдыньские диатомеи относятся к Бореальной палеобиогеографической области, а диатомеи Калифорнии и бассейна р. Апуки - к Индо-Тихоокеанской области.
Позднемеловые диатомеи являются самыми древними среди известных нам хорошо сохранившихся ископаемых диатомей. В отложениях кампана и Маастрихта найдены представители 59 родов и 290 видов, разновидностей и форм. До сих пор в отложениях мелового времени не обнаружены диатомеи с настоящим швом, но формы с осевым полем уже существовали. Морфологическое строение и тонкая структура панциря позднемеловых диатомей близки, а во многом и сходны с таковыми современных диатомей.
Роды и виды, составляющие флору диатомей кампана, имели различную длительность существования. Из 33 родов диатомей кампана 16 известны и в настоящее время, хотя многие представлены иными видами. Из 145 видов около 50% пока найдены только в отложениях позднего мела, остальные перешли в состав палеоценовой флоры, некоторые доживают до эоцена, олигоцена и даже неогена, единичные из меловых видов существуют и в современных морях.
Большое систематическое разнообразие диатомей мелового периода и их сложное морфологическое строение указывают на длительный пред-шествующий путь эволюционного развития. Истоки диатомей следует искать в более древних породах, и в первую очередь в отложениях юры и раннего мезозоя.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'