Строение и размножение харофитов
Строение клеток. Все клетки харовых водорослей в начальных стадиях дифференцировки отдельных частей таллома одноядерны (рис. 262, 1). При размножении этих клеток их ядро делится митотически (рис. 262, 2). Количества выявляющихся при этом хромосом у разных видов различное, от 6 до 70, так же как различны их размеры и форма (рис. 262, 3, 4). Только при последующей дифференцировке, когда определенные клетки вытягиваются и становятся междоузлиями или другими удлиненными частями таллома, в них происходят многократные амитотические деления ядра без образования клеточных перегородок, в результате клетки становятся многоядерными, а ядра - вытянутыми, неправильной формы (рис. 262, 5-7).
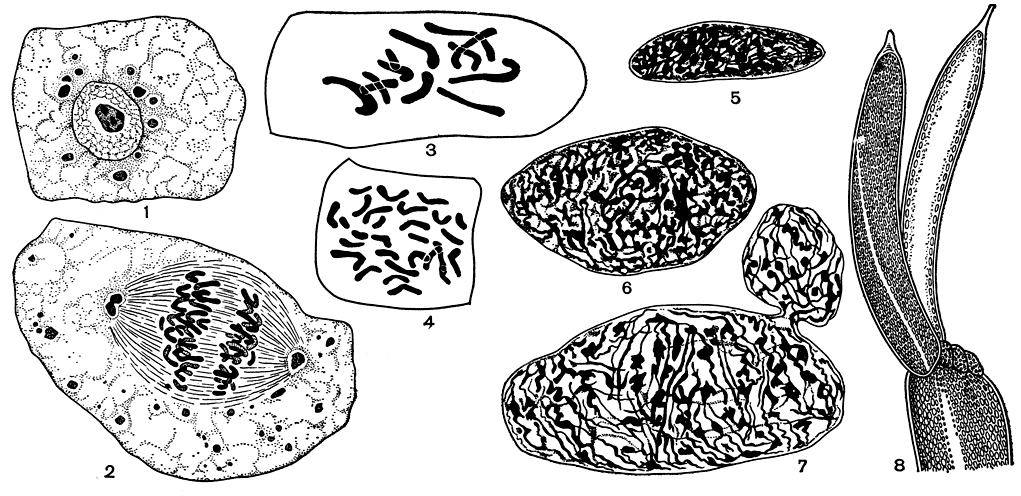
Рис. 262. Строение клеток харофитов: 1 - одна из клеток талломного узла с ядром в покоящемся состоянии (после специальной окраски, выявляющей его строение) (Х 1800); 2 - молодая клетка из растущей части таллома в стадии митотического деления ядра (после такой же окраски) (Х 2400); 3, 4- число (n), размеры и форма хромосом у разных представителей харофитов: 3 - n = 12, 4 - n = 28 (Х 1900); 5-7 - стадии формирования и последующего амитотического деления ядра из клетки, образующей талломное междоузлие (Х 1800); 8 - концевая часть листа нителлы: левый членик - вид с поверхности, правый членик - вид в продольном разрезе
Хлоропласты в клетках многочисленны и имеют форму мелких дисковидных телец, подобных хлорофилльным зернам высших растений. Набор ассимиляционных пигментов сходен с таковым у зеленых водорослей (хлорофиллы а и b и почти полностью те же каротиноиды), а в качестве запасного питательного вещества также вырабатывается крахмал.
Вытянутые сформировавшиеся клетки харофитов имеют очень характерное строение (рис. 262, 8). Их оболочка плотная, довольно толстая. Внутренний ее слой образован целлюлозой; наружный слой состоит из каллозы, в которой может отлагаться углекислая известь. Всю центральную часть клетки занимает вакуоля клеточного сока, а протоплазма распределена по периферии у внутренней поверхности оболочки. В тонком, непосредственно примыкающем к оболочке слое протоплазмы сосредоточены хлоропласты; в более толстом внутреннем слое, граничащем с вакуолью, расположены многочисленные ядра (рис. 262, 8, справа). Оба слоя протоплазмы, если рассматривать клетку под микроскопом, показывают ряд интересных особенностей. При установке фокуса микроскопа на поверхность клетки (рис. 262, 8, слева) прежде всего бросается в глаза, что хлоропласты распределяются равномерно, окрашивая всю клетку в сплошной ярко-зеленый цвет. Исключение составляет только узкая бесцветная полоса, идущая вдоль всей клетки чуть наискось - она лишена хлоропластов, но определяет их положение в остальных частях клетки: хлоропласты расположены ровными рядами параллельно этой бесцветной полосе. Если сфокусировать микроскоп на внутренний слой протоплазматического мешка, то при достаточно большом увеличении в живых клетках открывается удивительная картина стремительного движения протоплазмы по всей клетке, параллельно той же продольной бесцветной линии: по одну ее сторону в одном на-правлении, по другую сторону в прямо противоположном направлении. Движение протоплазмы в клетках, как известно, свойственно всем растениям, но у харовых водорослей оно отличается наибольшей скоростью, равной 1,5-2 мм в минуту, что при сильных увеличениях микроскопа производит впечатление значительной быстроты.
Строение верхушки основных побегов. Сложность строения таллома харовых водорослей легче всего понять, если проследить дифферен-циацию клеток в процессе верхушечного роста основного побега (стебля). В простейшей форме это выражено у тех харофитов, талломы которых не покрыты особым слоем клеток, именуемым корой, например у видов нителлы.
Верхушечная клетка стебля имеет характерную плоско-выпуклую форму, т. е. нижняя перегородка, отделяющая ее от нижележащей клетки, плоская, а верхняя свободная стенка полусферическая (рис. 263, 1). В течение вегетационного периода верхушечная клетка непрерывно делится горизонтальной (перпендикулярной к длинной оси стебля) перегородкой на две клетки, из которых верхняя остается верхушечной полусферической, нижняя становится цилиндрической. При следующем делении этих двух клеток такими же горизонтальными перегородками получается комплекс из четырех клеток, расположенных друг за другом, который является исходным для всего последующего процесса дифференциации и роста таллома. Новое деление верхушечной клетки повторяет описанный процесс возникновения цилиндрической клетки, а прежняя цилиндрическая клетка делится на верхнюю двояковогнутую клетку и нижнюю двояковыпуклую клетку (рис. 263, 2). Дальнейшая судьба этих последних двух клеток резко различна: верхняя клетка делится и становится исходной для формирования многоклеточного узла (рис. 263, 3а), тогда как нижняя оказывается лишенной способности делиться и дальше только растет в длину, образуя междоузлие (рис. 263, 3 б).
Деление исходной узловой клетки происходит уже не горизонтальными, а вертикальными (параллельными оси стебля) перегородками, которые располагаются таким образом, что после ряда делений образуются две клетки в центре узла и несколько клеток по его периферии, что хорошо видно на поперечном срезе через узел (рис. 263, 4). В дальнейшем центральные клетки более уже не делятся, а периферические продолжают делиться и дают начало листьям и боковым ветвям стебля (а у некоторых представителей еще так называемым прилистникам и коре).
Из каждой периферической клетки стеблевого узла возникает по одному листу. Листья закладываются первоначально как бугорки (рис. 263, 5 в), которые лишь постепенно вытягиваются и развиваются в листья. При этом периферические узловые клетки стебля выполняют роль верхушечных ростовых клеток, подобных верхушечным клеткам стебля. Однако, в отличие от последних, они делятся не беспредельно, а отчленяют от себя только определенное количество сегментов. Эти сегменты, разрастаясь и дифференцируясь, образуют членистые листья, состоящие из длинных неделящихся клеток и чередующихся с ними узлов, сложенных несколькими мелкими клетками, способными к дальнейшим делениям. Один или несколько конечных члеников листа лишены узлов. Подобное строение делает листья принципиально сходными со стеблем, но функция узлов у них уже совершенно другая. Из периферических клеток большинства листовых узлов образуются органы полового размножения и окружающие их придаточные клетки. Только самый нижний, так называемый базальный узел, находящийся в основании каждого листа (рис. 263, 6 г), имеет другую функцию. Отдельные его клетки, расположенные в пазухе листьев, дают начало боковым ветвям стебля (рис. 263, 6д), другие служат в некоторых родах харофитов для образования так называемых прилистников и коры (рис. 263, 6е).
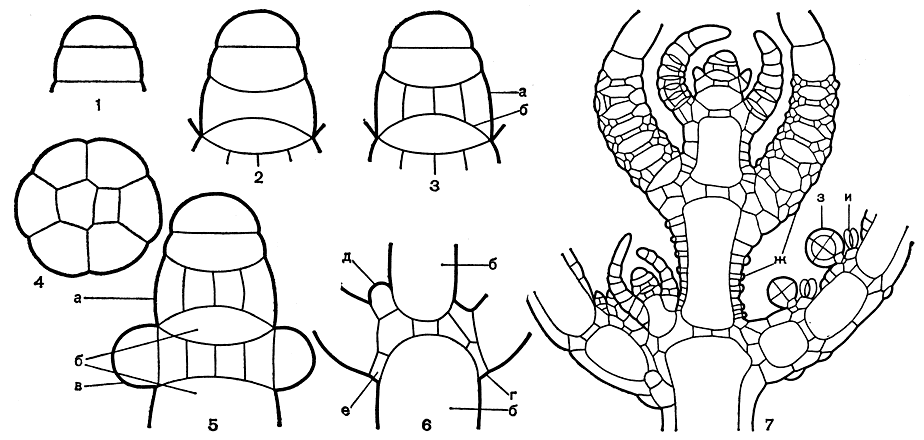
Рис. 263. Строение верхушки стебля харофитов в продольном сечении (схематизировано): 1-6 - схема начальных стадий верхушечного роста (1 - двухклеточная стадия, 2 - четырехклеточная стадия, 3 - начало формирования стеблевого узла и междоузлия, 4 - поперечный разрез сформированного стеблевого узла, 5 - начальная стадия образования листьев из периферических клеток стеблевого узла, 6 - стеблевой узел с развитыми листьями и образующейся ветвью); 7 - часть стебля хары на протяжении верхних четырех мутовок, а - стеблевой узел, б - междоузлие, в - начальные листовые бугорки, г - базальный листовой узел, д - начальный бугорок боковой ветви стебля, е - клетки, образующие прилистники, ж - клетки стеблевой и листовой коры, з - антеридии, и - оогонии
Прилистники. У всех видов хары и у представителей некоторых других родов харофитов на стеблях при основании листовых мутовок имеется венчик более или менее вытянутых и заостренных, а иногда и очень длинных или, наоборот, почти шаровидных клеток, получивших условное название прилистников. Они образуются из двух боковых клеток базального узла каждого листа независимо от того, имеется ли у данного вида стеблевая кора или нет. В зависимости от количества делений каждой такой начальной клетки и степени разрастания получающихся от этого клеток может возникнуть несколько вариантов строения прилистников, характерных для разных видов. Если делению подвергается только одна из этих двух клеток, и это деление однократно, а деление второй клетки подавлено, то на каждый лист возникает по одному прилистнику, располагающемуся между листьями (иногда они сдвигаются под лист). Такой венчик можно назвать однократным (редкий случай). Если так развиваются обе клетки по обе стороны листа, то венчик становится двукратным (частый случай). В обоих случаях возникает лишь один ряд прилистников, т. е. венчик прилистников может быть или однократным однорядным, или двукратным однорядным. Однако чаще всего встречается третий случай, когда каждая начальная клетка делится дважды и развивается еще второй нижний ряд прилистников, т. е. венчик оказывается двукратным двухрядным (рис. 264).
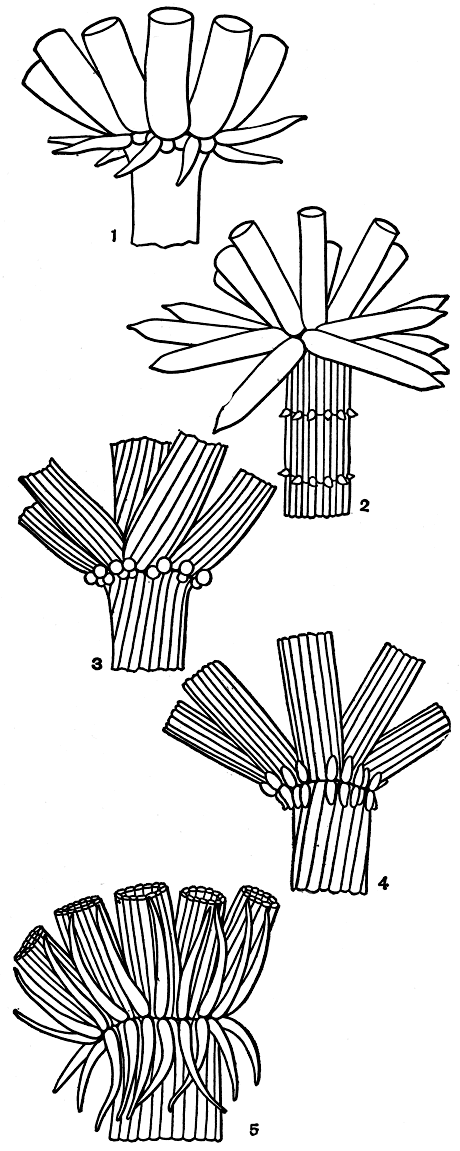
Рис. 264. Основания листовых мутовок разных видов хары с различно развитым венчиком прилистников: 1 - однократный однорядный венчик Chara braunii; 2 - двукратный однорядный венчик Ch. gymnopitys; 3-5 - двукратные двухрядные венчики (3 - Ch. fragilis, 4 - Ch. vulgaris, 5 - Ch. hispida)
Строение коры. Междоузлия на стеблях многих харофитов не остаются голыми клетками, как это характерно, например, для рода нителла, а обрастают слоем специальных клеток, получившим название коры. Стеблевая кора лучше всего выражена в роде хара, большинство видов которого снабжены ею. Стеблевая коpa харофитов всегда построена из полос клеток, расположенных параллельными рядами вдоль междоузлия слегка наискосок. Эти полосы бывают двух сортов - первичные, составляющие основу коры, и вторичные, усложняющие ее структуру, но эти последние возникают не у всех видов. Количество первичных полос соответствует количеству листьев в мутовке, что позволяет понять их происхождение.
От основания каждого листа отходят две первичные коровые полосы: одна - вниз по стеблю, другая - вверх. Их образуют две периферические клетки базального листового узла, расположенные посередине: одна - под листом, другая - над ним. Каждая из этих клеток выполняет функцию верхушечной ростовой клетки, она делится и развивается так же, как это было описано для роста стебля. В итоге каждая первичная коровая полоса оказывается состоящей из верхушечной клетки и чередующихся узлов и междоузлий. Узловые клетки небольшие, округлые, а клетки междоузлий крупные, вытянутые, более или менее цилиндрические. По мере роста первичные коровые полосы, идущие вниз по стеблю от листьев одной мутовки, примерно в середине стеблевого междоузлия встречаются с другими первичными коровыми полосами, идущими вверх от нижележащей мутовки. Таким образом, количество первичных коровых полос всегда равно количеству образовавших их листьев в соответствующей мутовке. Когда встречные клетки смыкаются, рост первичных коровых полос заканчивается. Волнистая линия смыкания коровых полос бывает хорошо заметна посередине стеблевых междоузлий. Кора, составленная только из одних первичных полос, получила название однополосной (рис. 265, 1). Она встречается у определенной, сравнительно небольшой группы видов рода хара.
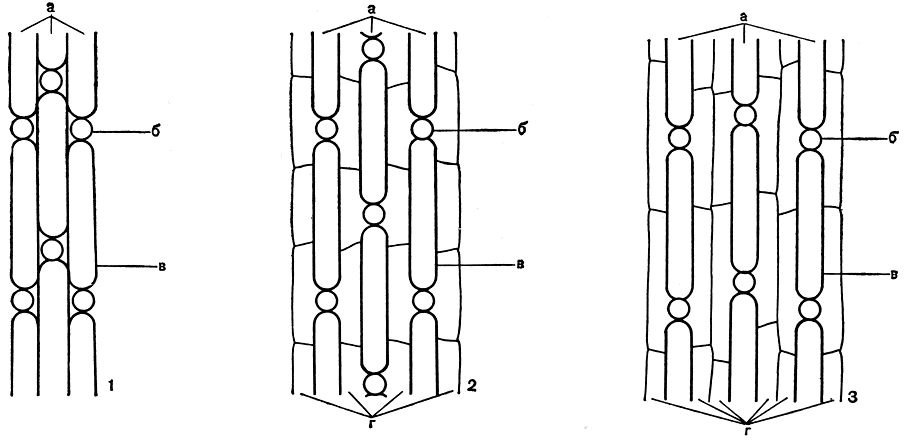
Рис. 265. Схемы строения основных типов стеблевой коры в роде хара (вид с поверхности): 1 - однополосная кора; 2 - двухполосная кора; 3 - трехполосная кора, а - первичные коровые полосы, б - их узлы, в - их междоузлия, г - вторичные коровые полосы
Строение такой коры усложняется в силу того, что узловые клетки первичных коровых полос способны делиться в двух направлениях. Если плоскости деления располагаются фронтально (параллельно поверхности стебля), то это приводит к образованию так называемых шипов - обычно вытянутых и заостренных клеток, одиночных или по 2-5 в пучках, направленных более или менее перпендикулярно к стеблю. Такие шипы в широкой мере свойственны однополосной коре (рис. 266, 1).
Второе, боковое направление делений узловой клетки коры имеет совершенно другие последствия - появление по обе ее стороны двух клеток, разрастающихся по стеблю в длину в промежутках между первичными коровыми полосами вверх и вниз каждая. В результате из этих двух клеток получается фигура в форме буквы Н с узловой клеткой в середине. Эти боковые клетки, образованные от всех узлов первичной коры, в процессе разрастания по стеблю смыкаются друг с другом и также слагаются в полосы, получившие название вторичных коровых полос. Однако здесь возможны два случая. У одних видов хары вторичные коровые клетки сравнительно короткие, равные примерно половине длины клеток коровых междоузлий, и тогда они могут образовать лишь одну вторичную коровую полосу между двумя первичными путем встречи концами противоположных (правых с левыми) клеток, образованных от узлов двух соседних первичных коровых полос (рис. 265, 2). Такую кору, когда между двумя первичными коровыми полосами располагается одна вторичная (на одну первичную приходится одна вторичная), называют двухполосной корой.
Наконец, если вторичные коровые клетки по длине равны клеткам междоузлий первичных коровых полос, то тогда происходит встреча концами однозначных клеток, образованных от узлов одной и той же первичной коровой полосы по обе ее стороны (правых с правыми, левых с левыми). В силу того что такие две вторичные полосы (правую и левую) образует каждая первичная коровая полоса, то по их боковым сторонам вдоль всего стеблевого междоузлия происходит смыкание двух параллельно идущих вторичных полос, происходящих от двух соседних первичных полос (рис. 265, 3). Кору такого строения, когда между двумя первичными коровыми полосами находятся две вторичные (на одну первичную приходятся две вторичные), называют трехполосной. Вторичные коровые полосы отличаются от первичных отсутствием округлых узловых клеток.
На узловых клетках двух- и трехполосной коры, как и в первом случае, могут образовываться шипы (рис. 266, 2-5). Видов хары с двух- и трехполосной стеблевой корой значительно больше, чем с однополосной. Еще малочисленное хары, вообще лишенные коры.
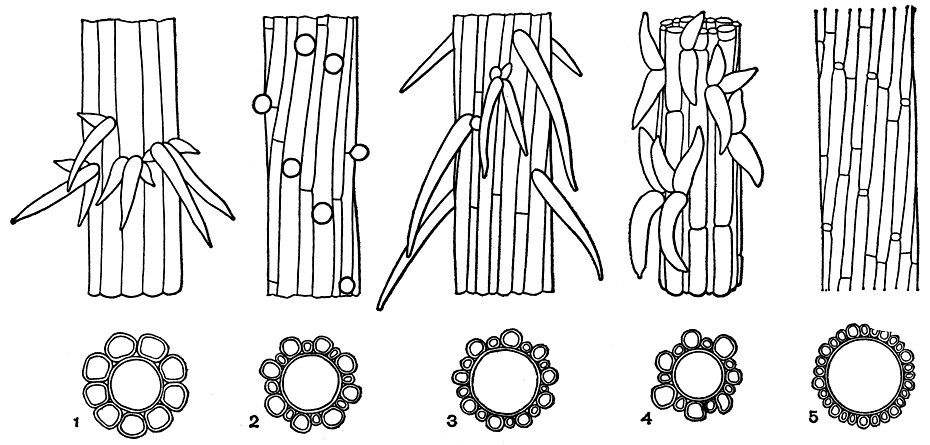
Рис. 266. Участки стебля разных видов хары с различно развитыми корой и шипами (вид с поверхности и в поперечном разрезе): 1 - вид с однополосной корой (Chara canescens); 2-4 - виды с двухполосной корой: 2 - первичные коровые полосы крупнее вторичных, шипы рудиментарные (Ch. contraria), 3 - то же, шипы длинные (Ch. aculeolata), 4 - первичные коровые полосы мельче вторичных (Ch. rudis); 5 - вид с трехполосной корой (Ch. fragilis)
На листьях хар кора образуется сходным образом - путем встречного нарастания коровых полос из листовых узлов вверх и вниз, и только самый нижний членик листа имеет одни нисходящие коровые полосы, идущие от первого листового узла вниз, так как базальный листовой узел восходящих коровых полос не образует. Однако строение коровых полос очень упрощено - каждая коровая полоса на всем протяжении листового междоузлия состоит только из двух одинаковых цилиндрических клеток, выросших из двух соседних узлов и встретившихся в середине листового междоузлия.
Листовая кора может быть только в том случае, если стебель покрыт корой. Однако у разных видов хары встречаются разные варианты развития коры. У одних при покрытом корой стебле листья могут все же оставаться голыми, без коры; у других с корой бывают или все членики листа, или только несколько первых члеников при длинных голых концах, образованных рядом простых клеток. Бывает, что без коры остается лишь первый членик листа.
Образование и строение антеридиев и оогониев. Антеридии и оогонии образуются из особых клеток, отчленяемых некоторыми периферическими клетками листовых узлов. Они могут возникать или вместе на одном талломе (однодомные виды), или раздельно на разных талломах (двудомные виды).
Формирование антеридия легко проследить у нителлы. Начальная клетка, отчлененная от узловой, делится на две, из которых наружная служит для образования собственно антеридия и быстро делится в трех взаимно перпендикулярных плоскостях на восемь клеток, слагающихся в шар, а прилегающая к листовому узлу является исходной клеткой, формирующей основание антеридия (рис. 267, 1). При дальнейшем делении клеток в верхней шаровидной части происходит прежде всего обособление восьми наружных клеток, которые далее уже не делятся, и восьми внутренних (рис. 267, 2), разделяющихся еще раз продольными перегородками, в итоге чего будущий антеридий оказывается состоящим из восьми наружных, восьми средних и восьми внутренних клеток, плотно соприкасающихся друг с другом (рис. 267, 3). Одновременно исходная клетка основания делится горизонтальной перегородкой на наружную базальную клетку, остающуюся далее без особых изменений, и внутреннюю базальную, так называемую бутылковидную клетку, сильно разрастающуюся впоследствии и вдающуюся внутрь антеридия, образуя его ножку (рис. 267, 2, 3). Последующий процесс дифференциации приводит к формированию восьми плоских покровных клеток треугольной или четырехугольной формы с характерными лучистыми утолщениями оболочки, смыкающихся своими зазубренными краями. Эти клетки, называемые щитками, образуют наружную стенку антеридия. Примыкающие к ним изнутри клетки среднего ряда постепенно вытягиваются и превращаются в так называемые рукоятки, несущие на конце восемь округлых головок, образованных восемью внутренними клетками (рис. 267, 4). При созревании антеридия эти основные головки образуют обычно по 6 вторичных головок каждая, и уже на них развиваются длинные антеридиальные нити, обычно по 4 на головке, состоящие примерно из 200 расположенных в один ряд клеток. В каждой клетке образуется по одному длинному спиралевидно изогнутому антерозоиду, с двумя жгутами на переднем конце (рис. 267, 6-9). Таким образом, каждый антеридий дает до 40 000 антерозоидов.
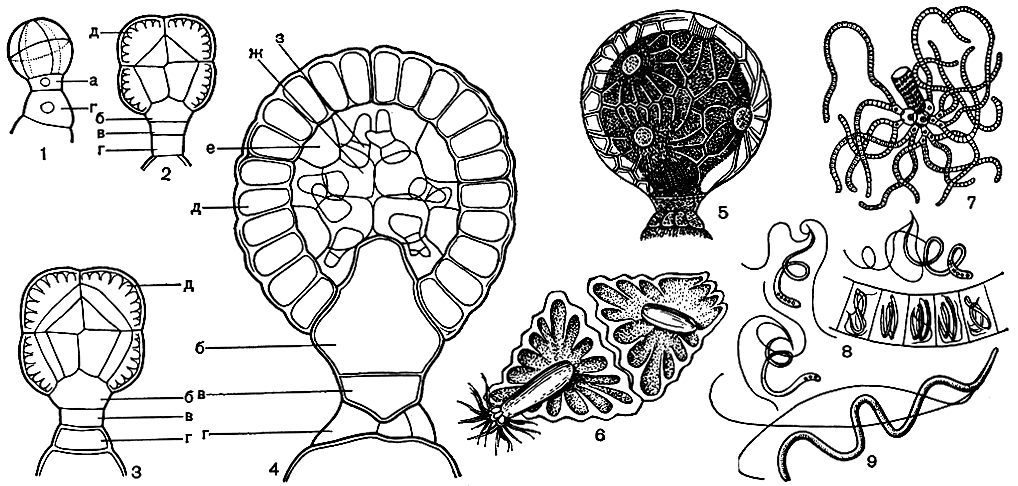
Рис. 267. Строение антеридия харофитов: 1-4 - последовательные стадии образования антеридия, вид с поверхности (1) и в продольном разрезе (2-4): 1 - начальная восьмиклеточная стадия, 2, 3 - последующие стадии обособления наружных щитовых клеток и внутренней части, 4 - поздняя стадия с дифференцированной внутренней частью; 5 - зрелый антеридий, вид с поверхности; 6 - два щитка с рукоятками, вид изнутри; 7 - рукоятка отдельно с хорошо развитыми головками и антеридиальными нитями; 8 - участок антеридиальной нити с антерозоидами в клетках и снаружи; 9 - антерозоид при большом увеличении, а - исходная клетка основания, б - внутренняя базальная ('бутылковидная') клетка, в - наружная базальная клетка, г - клетки листового узла, д - стенка, е - рукоятка, ж - головки, з - антеридиальные нити
Зрелые антеридии у всех харофитов имеют вид шарика на короткой ножке, диаметром в среднем около 0,5 мм, характерно пересеченного крест-накрест зигзагообразной линией смыкания щитков, в каждом из которых хорошо заметны по краям радиальные утолщения оболочки и в центре округлое основание внутренней рукоятки (рис. 267, 5). Антеридии сначала окрашены в зеленый цвет от содержащихся в щитках хлоропластов, затем они становятся кирпично-красными.
Формирование оогониев происходит значительно проще. Начало этому процессу кладет одна из клеток листового узла, составляющая в дальнейшем ножку оогония. Вверх она отчленяет одну центральную клетку, вбок вокруг себя - пять будущих обволакивающих клеток (рис. 268, 1). Центральная клетка делится один или два раза, в результате возникают неравновеликие клетки: крупная клетка, развивающаяся далее в яйцеклетку, и одна - три мелкие стерильные клетки, получившие название "поворотных", так как они, возникнув сбоку или сверху от яйцеклетки, постепенно перемещаются к ее основанию. Их считают сильно редуцированным остатком стенки собственно гаметангия. Одновременно с этим пять боковых клеток разрастаются вокруг яйцеклетки вверх по спирали справа налево и полностью обволакивают ее. На самой вершине каждая из них отчленяет одну или две небольшие клеточки, слагающие так называемую коронку (рис. 268, 2-4).
Зрелые оогонии имеют вид эллипсоидных или яйцевидных образований длиной до 1 мм, спиралевидно-исчерченных обволакивающими клетками и увенчанных небольшой или довольно крупной коронкой. Коронка из десяти мелких клеток, расположенных в два яруса, характерна для нителловых (рис. 268, 5); одноярусная коронка из пяти более крупных клеток - для собственно харовых (рис. 268, 6).
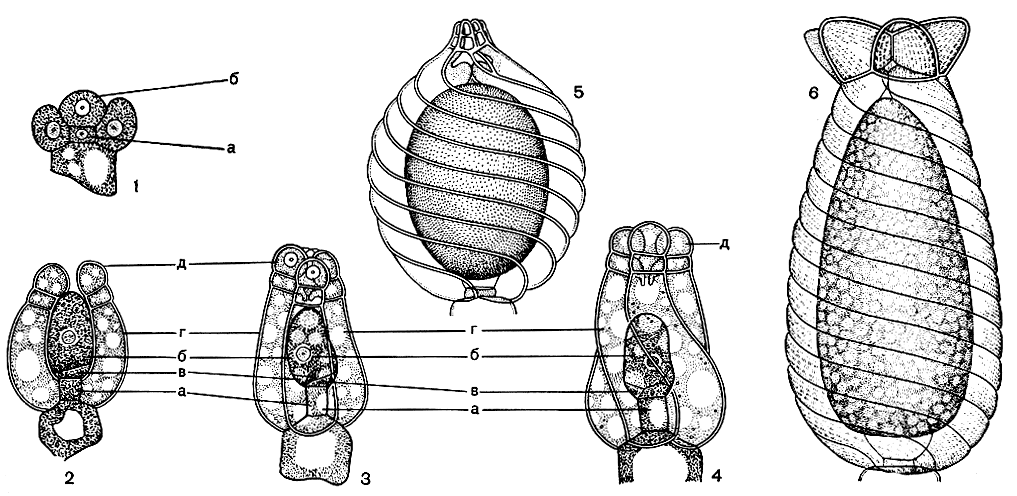
Рис. 268. Строение оогония харофитов: 1-4 - последовательные стадии образования оогония (1- начальная стадия, 2-4 - последующие стадии формирования яйцеклетки и обволакивающих спиральных клеток); 5 - зрелый оогоний нителлы (Nitella); 6 - зрелый оогоний хары (Chara). а - клетка листового узла, б - яйцеклетка, в - поворотные клетки, г - обволакивающие клетки, д - клетки коронки
Ооспоры и их прорастание. Ооспоры, развивающиеся внутри оогониев в результате оплодотворения яйцеклетки, имеют у разных видов различную форму и размеры, от почти шаровидных до узкоэллипсоидных, от очень мелких, длиной около 0,3 мм, до крупных, почти в 1 мм. По мере созревания наружная стенка ооспор затвердевает, пропитывается суберином и кремнеземом и окрашивается. Зрелые ооспоры бывают коричневато-желтыми, коричневыми или темно-коричневыми до черных, снаружи они всегда слегка блестящие. Стенки их образованы четырьмя оболочками - двумя окрашенными и двумя бесцветными. Наиболее прочной и наиболее интенсивно окрашенной является самая наружная оболочка, которая образуется из плотно прилегающих к ооспоре внутренних частей обволакивающих клеток оогония, претерпевающих соответствующие изменения. Благодаря такому происхождению этой оболочки ооспоры на поверхности несут спирально извитые ребра, представляющие собою слепки швов соприкосновения спиралевидных обволакивающих клеток оогония. Соответственно этому ребра идут от углов пятигранного основания ооспоры вверх справа налево. Особенно сильно развиты ребра на ооспорах у представителей семейства нителловых, где они имеют вид широкой бахромы, слагающей наверху ооспоры подобие высокого воротничка (рис. 269, 2). В семействе собственно харовых ребра на ооспоpax обычно невысокие, часто едва выдающиеся над поверхностью; наверху ооспоры они часто собираются в небольшой хохолок, а внизу переходят от каждого угла пятигранного основания в длинные коготки (рис. 269, 2).
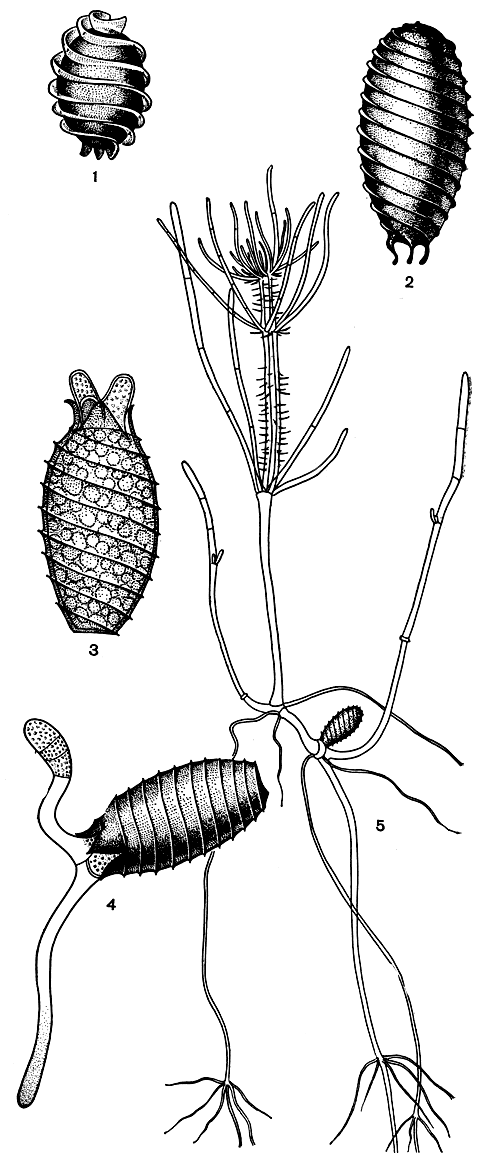
Рис. 269. Ооспоры и их прорастание: 1, 2 - внешний вид зрелых ооспорнителлы (1) и хары (2) 3-5 - последовательные стадии прорастания ооспор и формирования молодого растения хары (Chara canescens)
Зрелые ооспоры у многих видов покрываются еще слоем извести; внутри они заполняются запасными питательными веществами, главным образом зернами крахмала и каплями жира.
После периода покоя ооспоры прорастают. Прорастание начинается с того, что в верхней части ооспоры грубые крахмальные зерна и капли жира постепенно исчезают, заменяясь зернистой протоплазмой, а находящееся здесь ядро, ставшее после оплодотворения яйцеклетки диплоидным, претерпевает сперва одно деление, сопровождаемое мейозом, затем второе, в итоге чего получаются четыре гаплоидных ядра. Далее верхний участок протоплазмы с одним из этих ядер отделяется поперечной перегородкой, образуя линзообразную центральную клетку, от которой берет начало новое растение. Остальная часть ооспоры выполняет функции склада запасного питательного материала.
Одновременно с этими внутренними изменениями наружная твердая стенка ооспоры расщепляется на ее вершине на пять зубцов, давая выход проростку, возникающему из центральной клетки (рис. 269, 5). Первое деление центральной клетки происходит поперечной перегородкой, перпендикулярной к ее длинной оси, и приводит к образованию двух функционально различных клеток. Из одной, более крупной клетки в дальнейшем образуется стеблевой побег, который на начальной стадии развития называют предростком, из другой, меньшей клетки - первый ризоид. Оба они растут путем поперечных клеточных делений. Предросток растет вверх и довольно быстро зеленеет, заполняясь хлоропластами, первый ризоид направляется вниз и остается бесцветным (рис. 269, 4). После ряда клеточных делений, сообщающих им строение однорядных нитей, происходит их дифференцировка на узлы и междоузлия, и дальнейший их верхушечный рост протекает уже так, как было описано выше для стебля. Из узлов предростка возникают вторичные предростки, мутовки листьев и боковые ветви стебля, из узлов первого ризоида - вторичные ризоиды и их мутовчатые волоски. Таким путем и формируется таллом, состоящий из нескольких стеблевых побегов в верхней части и нескольких сложных ризоидов в нижней части (рис. 269, 5).
Нижняя часть стебля, ризоиды и клубеньки. Внешние различия между стеблевой и ризоидной частью таллома харофитов отчетливы только в том случае, если с самого начала стеблевая часть развивалась над грунтом, а ризоидная часть была целиком погружена в донный ил. При нарушении этого полное принципиальное сходство обеих частей становится очевидным. Так, участки стебля, погруженные в грунт, быстро укореняются, образуя из своих узлов ризоиды и проростки, а узлы на участках ризоидов, вышедших на поверхность грунта, быстро дают стеблевые побеги. Иными словами, из узловых клеток обеих частей таллома при взаимной перемене условий могут возникать вегетативным путем новые целые растения. Кроме того, у харофитов имеются специальные органы вегетативного размножения, так называемые корневые и стеблевые клубеньки.
Образование клубеньков более всего характерно для представителей рода хара. Клубеньки могут развиваться как на ризоидах, так и на нижних частях стебля, погруженных в грунт. Ризоидные клубеньки бывают многоклеточными (рис. 270, 1, 2) и одноклеточными (рис. 270, 3). В обоих случаях они являются специальными образованиями, заполненными крахмальными зернами и несущими на себе узловые клетки, способные к делению и последующему формированию вегетативных органов. Стеблевые клубеньки всегда многоклеточны и представляют собой стеблевые узлы с сильно редуцированными и видоизмененными листьями. У видов хары они неправильные, разные по величине и форме (рис. 270, 4), а у нителлопсиса отличаются исключительной правильностью и изящной формой звездочек, лучи которых соответствуют листьям мутовки (рис. 270, 5-7). Эти клубеньки также богаты крахмалом и очень легко прорастают в новые растения.
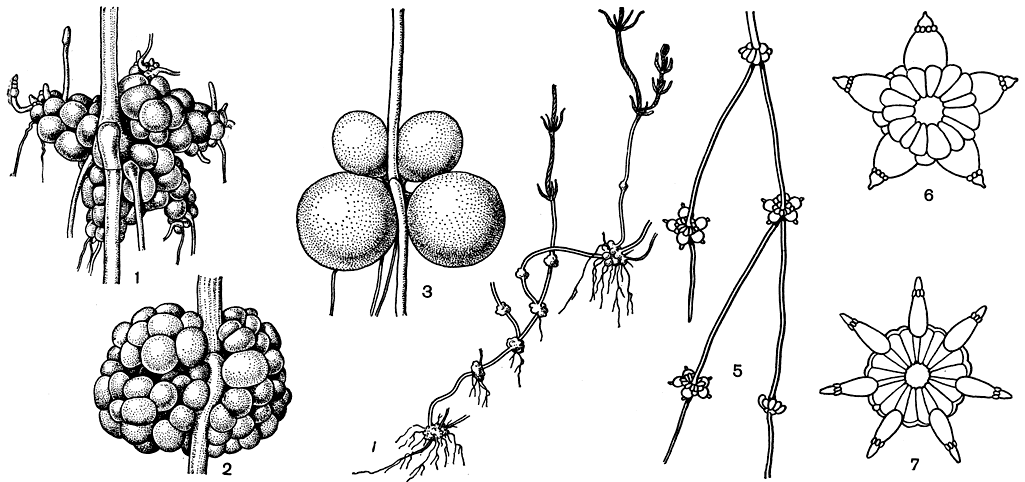
Рис. 270. Нижняя часть стебля, ризоиды и клубеньки харофитов: 1-3 - многоклеточные (1,2) и одноклеточные (3) корневые клубеньки хары, образовавшиеся на ризоидах (1 - Chara baltica, 2 - Ch. fragifera, 3 - Gh. aspera); 4-7 - клубеньки, образовавшиеся из стеблевых узлов на нижних погруженных в ил побегах: 4 - у Chara baltica (при слабом увеличении), 5-7 - у Nitellopsis obtusa (при слабом и сильном увеличении)
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'