Порядок сифоновые (Siphonales)
Слоевище водорослей этого порядка лишено перегородок, только в основании ветвей и изредка гаметангиев возникают своеобразные пробки. В пристенном слое цитоплазмы содержатся многочисленные ядра и хлоропласты с пиреноидами или без них.
Чтобы познакомиться со строением и биологией водорослей этого порядка, рассмотрим несколько наиболее характерных представителей трех различных семейств.
Семейство каулерповые (Caulerpaceae)
Многим известна водоросль каулерпа (Caulerpa) - типичный представитель семейства каулерповых (Caulerpaceae). Этот обширный род (около 60 видов) весьма характерен для флоры тропических морей. Лишь немногие виды заходят в субтропические широты, и среди них распространенная в Средиземном море каулерпа прорастающая (С. prolifera). Крупные стелющиеся растения каулерпы часто покрывают большие пространства морского дна. Поселяясь обычно на различных твердых грунтах - камнях, скалистых рифах, мертвых кораллах, каулерпа благодаря своему стелющемуся слоевищу и наличию многих ризоидов неплохо чувствует себя также на песчаных и илистых грунтах. Излюбленным местообитанием видов каулерпы является мелководье с более или менее спокойной водой, в удалении от постоянного прибоя, например лагуны, защищенные от действия прибоя коралловыми рифами.
Все каулерпы построены по одному плану. Ползучие, распростертые на грунте части слоевища, или ризомы, имеют вид цилиндрических сифонов. Ризомы весьма напоминают столоны некоторых наземных растений. Они тянутся в длину на десятки сантиметров, разветвляются и отчленяют через определенные интервалы вниз ризоиды и вверх вертикальные побеги. Ризоидов много, на конце они сильно и многократно ветвятся и тем самым обеспечивают плотное прикрепление растения к любому субстрату. Вертикальные побеги выполняют роль ассимиляторов. В них сосредоточены хлоропласты и происходит процесс фотосинтеза. Если ризомы и ризоиды у разных видов почти не отличаются по морфологии, то строение ассимиляционных ветвей достигает большого разнообразия и сложности и служит хорошим признаком для разграничения видов (рис. 229, 1-4). Они могут быть цилиндрическими, подобно ризомам, или уплощенными до листовидных. У большинства видов они снабжены многочисленными боковыми выростами разнообразной формы - серповидными, щитковидными, кеглевидными, метельчатыми и т. п.
Специфическим свойством каулерповых является образование внутреннего скелета. Во всех частях растения через полость сифона проходят особые переплетающиеся тяжи, которые ориентированы радиально или перпендикулярно поверхности (рис. 229, 5). Это отчетливо видно на поперечных срезах через слоевище. Они образуются в цитоплазме близ вершины ветвей. Первоначально они могут быть свободными на обоих концах, но нередко уже в момент возникновения связаны со стенками слоевища. Очень тонкие вначале, они быстро утолщаются и становятся слоистыми подобно общей оболочке растений. Функция внутреннего скелета остается неясной. Не исключено, что он играет какую-то роль в минеральном питании каулерпы. Стенки слоевища состоят из каллозы, кроме того, в них можно обнаружить пектиновые вещества. Отсутствие целлюлозы отличает каулерпу, как и все семейство каулерповых, от остальных водорослей порядка.
Кроме зеленых хлоропластов, для каулерповых характерно наличие бесцветных пластид, или лейкопластов, которые участвуют в процессе накопления крахмала. У всех других водорослей порядка найдены только хлоропласты.
Вегетативное размножение занимает заметное место в жизни этих водорослей: отломившиеся части растений, попадая в новые места, легко прикрепляются к субстрату и разрастаются.
Бесполое размножение у каулерповых отсутствует, по-видимому, оно было утрачено у современных форм в процессе эволюции. Половое размножение - анизогамия - характеризуется рядом особенностей, отличающих каулерповых от других семейств порядка. Во-первых, у каулерпы нет специальных органов размножения - гаметангиев. Гаметы образуются непосредственно в ассимиляционных нитях в любой их части. На отдельных участках цитоплазма сгущается, приобретает темно-зеленую окраску, затем сетчатое строение и наконец делится с образованием одноядерных гамет. Какие-либо перегородки, отделяющие место образования гамет, отсутствуют. Такое размножение называют голокарпией. Для выхода гамет на поверхность слоевища образуются довольно длинные выросты - папиллы; гаметы освобождаются в результате разрыва оболочки на вершинах папилл.
Долгое время считали, что семейство состоит всего из одного рода каулерпа. Но теперь, когда подробнее изучено строение и размножение других сифоновых, установлено, что к нему принадлежит еще несколько родов, которые раньше относили к семейству кодиевых. Из них следует упомянуть роды удотея (Udotea) и халимеда (Halimeda).
В отличие от каулерпы вертикальные ветви у них тесно переплетаются (рис. 229, 6), в результате чего формируется псевдопаренхимное слоевище с определенной и довольно сложной морфологией.
Виды удотеи характеризуются вееровидным слоевищем различной формы (табл. 32, 1) с ризомами в основании. Представители тропических видов этого рода более или менее сильно пропитаны известью. Интересно, что в пределах рода удотея можно проследить переход от форм со свободными ветвями к формам псевдопаренхимного строения.

Таблица 32. Улотриксовые и сифоновые водоросли: 1 - удотея ( Udotea flabellata); 2 - ацетабулярия (Acetabularia peniculus); 3 - ульвария (Ulvaria obscura); 4 - хетоморфа (Chaetomorpha melagonium); 5 - пенициллус (Penicillus capitatus); 6 - кодиум (Codium fragile)
Наиболее сложным строением характеризуется халимеда, у которой слоевище состоит из множества вееровидных сегментов, сильно пропитанных известью (табл. 33, 2). Около 25 видов этого рода известны из тропических и субтропических морей. Многие из них играют большую роль в формировании коралловых рифов.
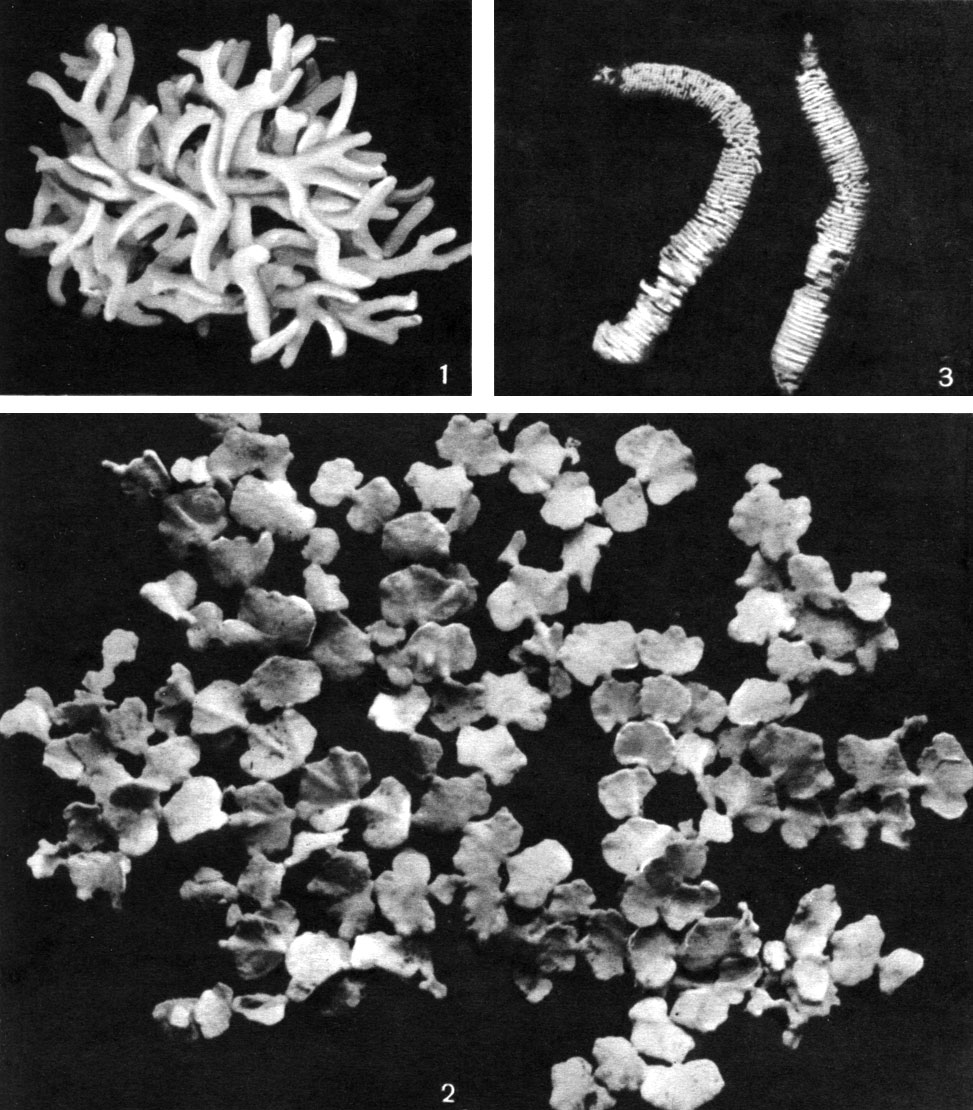
Таблица 33. Сифоновые водоросли: 1 - кодиум (Codium edule); 2 - халимеда (Halimeda opuntia); 3 - неомерис (Neomeris annulata)
Если рассмотреть ранние стадии развития халимеды и удотеи (рис. 229, 7-11), то бросается в глаза родство всех сифоновых водорослей. Вначале зигота растет, оставаясь какое-то время одноядерной. Затем, на более поздней стадии, она претерпевает ряд цитологических и морфологических изменений. Одно крупное ядро делится, некоторые хлоропласты теряют свой крахмал и превращаются в лейкопласты, вся структура вытягивается в вертикальный сифон с первичным ризоидом в основании. Следующая стадия имеет разнонитчатое строение. И только на самых поздних стадиях развития происходит переплетение и смыкание вертикальных нитей.
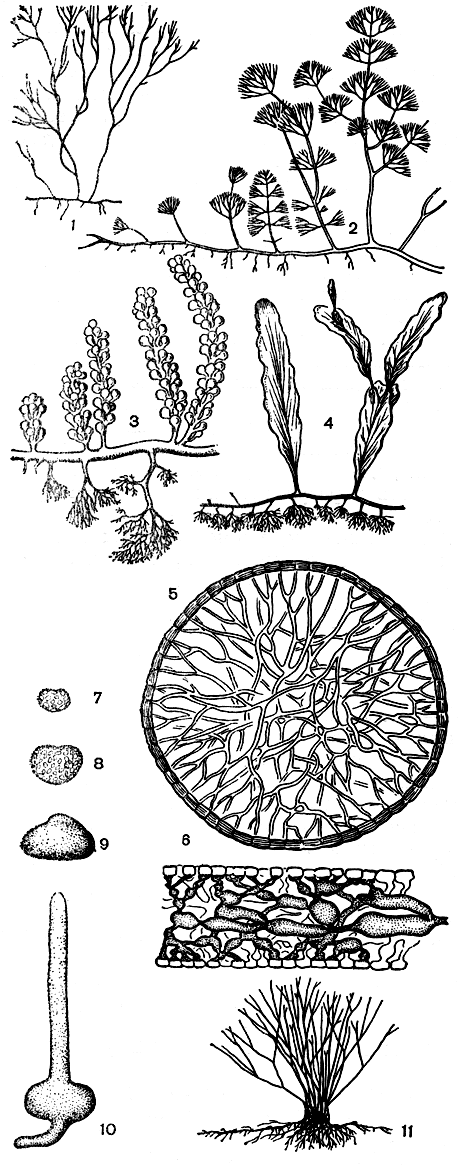
Рис. 229. Строение каулерповых: 1-5 - Caulerpa (1-4 - различные типы строения вертикальных ветвей, 5 - 'внутренний скелет' ризома на поперечном срезе); 6 - продольный срез слоевища Halimeda; 7-11 - ранние стадии развития Udotea
Семейство бриопсидовые (Bryopsidaceae)
Это семейство охватывает четыре рода, тесно связанные между собой.
Род дербезия (Derbesia) включает нитчатые водоросли, распространенные не только в тропических, но и в умеренных морях. Слоевище их состоит из стелющихся и вертикальных разветвленных нитей. В месте отхождения ветвей, так же как и в основании спорангиев, образуются своеобразные перегородки. Они могут быть простыми или двойными, и тогда между ними остается короткий сегмент с хлоропластами и ядрами. Вегетативное размножение, кроме фрагментации, заключается в образовании на вершине ветвей своеобразных выводковых почек, которые, отделяясь, превращаются в новые растения. Для бесполого размножения служат зооспоры (изредка апланоспоры). Это крупные шаровидные клетки с венцом жгутиков на переднем конце (рис. 230, 2-3), содержащие несколько ядер и хлоропластов. При их образовании обычные вегетативные веточки меняют свою форму на шаровидную или грушевидную, и, отделяясь от слоевища перегородкой, превращаются в спорангии.
Споры выходят из спорангиев через отверстие в их стенке и без периода покоя прорастают. Полового размножения нет.
Род халицистис (Halicystis) представлен несколькими видами, которые всегда растут на известковых красных водорослях типа литотамнион. Когда халицистис начинает прорастать, вначале образуются разветвленные нити, стелющиеся на поверхности субстрата. Позднее они проникают внутрь известковых водорослей, образуя короткие вертикальные побеги, на концах которых формируются пузыри диаметром до 3 см (рис. 230, 4-6). Таким образом, слоевище халицистиса не просто пузырь. В основе его строения лежит та же система разветвленных нитей, которая свойственна дербезии, так что принципиальной разницы между этими родами нет. Пузыри же есть не что иное, как видоизмененные вертикальные ветви. Их образование, вероятно, связано со "сверлящим" образом жизни этих водорослей.
Вегетативное размножение халицистисов происходит путем образования дополнительных пузырей на одном ризоме или нескольких новых пузырей на одном старом. Пузыри легко отделяются от слоевища, сохраняя при этом туpгop и способность к размножению.
Кроме вегетативного, известно и половое размножение - анизогамия. Гаметы образуются в верхней части пузырей в результате преобразования и деления участков цитоплазмы, которые отделяются от остальной части их лишь тонким цитоплазматическим слоем. Гаметы выходят под давлением через несколько отверстий. Один и тот же пузырь плодоносит обычно несколько раз. Бесполого размножения нет.
В настоящее время установлено, что халицистис и дербезия представляют собой разные формы развития одного жизненного цикла, причем размножающаяся зооспорами дербезия является спорофитом, а половые растения халицистиса - гаметофитами. Однако правильная смена гаметофита и спорофита у этих водорослей необязательна и каждая форма развития способна к длительному самостоятельному воспроизведению.
В природе известно намного больше видов дербезии, чем халицистиса; при этом первых можно встретить в тех районах, где вторые никогда не встречаются.
Род бриопсис (Bryopsis) также имеет спорофитную стадию типа дербезии. Основную часть слоевища бриопсиса составляет система вертикальных ветвей, тогда как стелющаяся часть развита в меньшей мере и присутствует в виде ризомов или всего лишь развитых ризоидов. В вертикальной части различается главная ось, покрытая ветвями первого и второго порядков. У одних видов слоевища перисто-разветвленные (рис. 230, 7), т. е. ветви у них располагаются супротивно и в одной плоскости; при этом верхние короче нижних. Все это делает растение похожим на перо птицы. У других ветви на слоевище располагаются вокруг всей оси радиально и тогда имеют метельчатое строение. В основании ветвей могут образовываться дополнительные ризоиды, которые в начальной стадии своего развития выглядят как короткие отростки, при сильном же развитии почти сплошь покрывают главную ось (рис. 230, 8). В месте образования ветвей наблюдаются перетяжки, ветви здесь сужаются и внутренняя часть оболочек утолщается. С поверхности эти утолщения можно принять за перегородки, на самом же деле они не отделяют ветвей, и все растение бриопсиса содержит одну центральную вакуолю, ограниченную пристенным слоем цитоплазмы.
Стелющиеся нити бриопсиса, или ризомы, служат для переживания неблагоприятных условий и могут быть многолетними, тогда как вертикальная часть существует только в определенный сезон. В основании вертикальных ветвей постоянно возникают новые ризомы, которые, в свою очередь, опять дают прямостоячие ветви. Благодаря этому бриопсис растет дерниной. Обычный способ вегетативного размножения заключается в том, что веточки отделяются от растений и укореняются при помощи ризоидов, образующихся в их основании.
Половой процесс - анизогамия, как и у других сифоновых. Гаметы образуются в конечных веточках, которые, перед тем как стать гаметангиями, не меняя формы, отделяются перегородкой от остального слоевища. Перегородки образуются как внутренние кольцеобразные утолщения боковой стенки в основании веточек. Эти утолщения, разбухая, оставляют в середине лишь узкий тяж протоплазмы, но постепенно и этот тяж прерывается, так как цитоплазма веточки и главной оси одевается собственной оболочкой, а первоначальное утолщение остается как студневидная пробка между этими оболочками. Выход гамет осуществляется в результате ослизнения и растворения оболочки на вершине веточки или через боковое отверстие в боковой стенке.
Таким образом, у бриопсиса мы видим уже специализированные органы полового размножения, но, поскольку они сохраняют ту же морфологию, что и вегетативные веточки, можно считать, что дифференциация гаметангиев здесь еще не закончена.
Зигота прорастает без периода покоя. Во многих случаях при прорастании вновь формируются растения бриопсиса, размножающиеся половым путем, но у некоторых видов обнаружена смена форм развития. У них зигота прорастает в нитчатое слоевище, ничем не отличающееся от дербезии и размножающееся сходными зооспорами. Зооспоры, в свою очередь, развиваются в половые растения бриопсиса. Иными словами, в этой группе водорослей наблюдается интересное явление. Разные роды - халицистис и бриопсис - имеют одну и ту же спорофитную стадию дербезии, с той лишь разницей, что у халицистиса смена гаметофита и спорофита происходит более регулярно и часто, тогда как у бриопсиса это случается реже и большинство видов этого рода размножается только половым путем.
Род псевдобриопсис (Pseudobryopsis) обладает наиболее высокой организацией в семействе. По строению и внешнему виду он очень сходен с бриопсисом, но отличается наличием перегородок в основании вегетативных веточек и более дифференцированными гаметангиями. Веточки, в которых образуются гаметы, видоизменяются, приобретая урновидную или грушевидную форму. Освобождаются гаметы через верхушечную пору (рис. 230, 11).
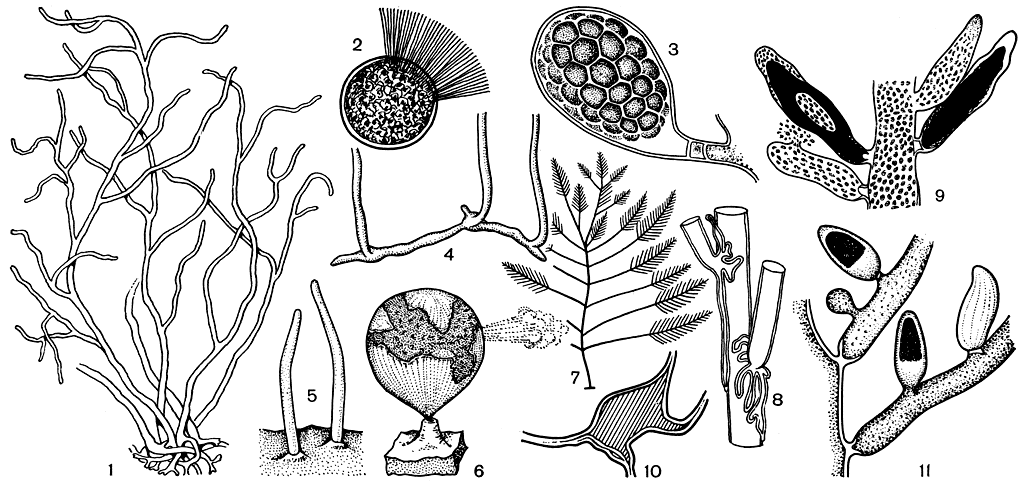
Рис. 230. Строение бриопсидовых: 1-3 - Derbesia (1 - внешний вид слоевища, 2 - зооспора, 3 - зооспорангий); 4-6 - Halieystis (4 - первичные нити, 5 - молодые вертикальные нити, 6 - зрелый пузырь); 7-10 - Bryopsis (7 - веточка, 8 - ризоиды в основании веточек, 9 - часть слоевища с гаметангиями, 10 - перегородка в основании гаметангия в продольном сечении); 11 -часть слоевища Pseudobriopsis с гаметангиями
Семейство кодиевые (Codiaceae)
Семейство кодиевых содержит только один род кодиум (Codium), насчитывающий в настоящее время около 50 видов (табл. 31,4; 32, 6; 33, 1). Будучи характерными для флоры субтропических морей, они встречаются и в умеренных широтах, проникая в холодные воды северного и южного полушарий. Кодиум хрупкий (С. fragile) найден в Норвегии; другой вид - кодиум Риттера (С. ritteri) - обильно развивается на Командорских островах. В южном полушарии кодиумы доходят до субантарктических районов. Чаще всего их можно встретить на глубине 50-230 м и глубже, но некоторые виды выходят на литораль. Они предпочитают чистые воды и поселяются на различных твердых грунтах.
Слоевище кодиума обычно небольших размеров, длиной 5-30 см, но есть крупные виды, как, например, кодиум большой (С. magnum), длиной до 8 м. Темно-зеленое губчатое слоевище имеет псевдопаренхимное строение. Оно состоит из множества более или менее плотно переплетенных неклеточных нитей. По форме слоевищ можно выделить две группы видов. Одни имеют распростертое слоевище, шаровидное или подушковидное; другие - вертикальное, цилиндрическое или сдавленное, обильно разветвленное. Для прикрепления растений служат ризоиды.
Принцип анатомического строения кодиума легко понять, глядя на продольный срез через слоевище (рис. 231, 1). В нем можно различить две зоны. Внутренняя образована пучком относительно тонких, бесцветных, более или менее густо разветвленных нитей. От нитей, расположенных по периферии пучка, отчленяются широкие и короткие пузыри, или утрикулы. Они располагаются таким образом, что окружают внутренний пучок бесцветных нитей, образуя поверхностный, или коровой, слой слоевища. Пузыри окрашены и выполняют ассимиляционную роль. Если внутренние нити у всех видов весьма сходны, то строение пузырей (их форма, размеры, очертания оболочек на вершине и т. п.) различно и служит одним из основных видовых признаков. В верхней части пузырей отходят бесцветные, длинные, легко отпадающие волоски, значение которых неясно. Если волоски развиваются обильно, все растение приобретает опушенный вид.
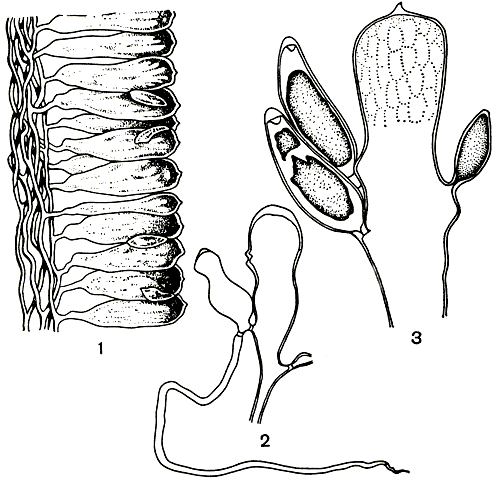
Рис. 231. Строение кодиума (Codium): 1 - часть продольного среза через слоевище; 2 - пузырь с дополнительным пузырьком; 3 - пузырь с гаметангиями
Рост слоевища в длину обусловлен апикальным вытягиванием центральных нитей. Образование новых пузырей среди уже существующих не только ведет к более плотному их расположению, но и представляет собой форму интеркалярного роста слоевища. У стелю-щихся подушковидных форм этот способ роста преобладает.
Многочисленные мелкие хлоропласты лишены пиреноидов. Лейкопласты отсутствуют. Оболочки кодиума состоят из каллозопектиновых веществ, но, в отличие от каулерповых, в них обнаружено небольшое количество целлюлозы.
Известно, что кодиум размножается вегетативным и половым путем. Вегетативное размножение широко распространено и происходит посредством фрагментации или образования нескольких слоевищ от одного основания. Кроме того, для кодиума характерно образование специальных дополнительных пузырьков с ризоидом в основании, которые, отделяясь и укореняясь, дают начало новым растениям. Бесполое размножение неизвестно; вероятно, у этого рода оно полностью утеряно в процессе эволюции.
Гаметы у кодиума двужгутиковые, грушевидные. Женские гаметы содержат многочисленные хлоропласты с пиреноидами; мужские - всего 1-3 хлоропласта, лишенные пиреноидов. Половой процесс, как правило, анизогамия. Дифференцированные гаметангии образуются как боковые выросты грушевидной формы на пузырях и отделяются от них специальными слизистыми перегородками, как у бриопсиса. Когда гаметы созревают, оболочка на вершине гаметангия разрывается, и гаметы выходят в окружающую среду. Зигота прорастает обычно без периода покоя. Новое поколение опять дает гаметы, и цикл развития завершается.
Изучение рода кодиум показывает, что семейство кодиевых, будучи очень близким к бриопсидовым, представляет по сравнению с ними более высокоорганизованную и специализированную группу. Более высокая организация проявляется в псевдопаренхимном строении, в наличии дифференцированных органов размножения и в полной утрате бесполого размножения. В настоящее время бриопсидовые и кодиевые рассматриваются как отдельные семейства, но их отличия говорят лишь о большей примитивности первых и эволюционной продвинутости вторых, принципиальных же различий между ними нет. Оба семейства представляют собой единую в пределах порядка ветвь эволюции, отличную от каулерповых и, по-видимому, более молодую по сравнению с ними.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'