Порядок родимениевые (Rhodymeniales)
Родимениевые - небольшой, достаточно хорошо отграниченный порядок, занимающий обособленное систематическое положение в классе флоридеевых. Основное отличительное свойство родимениевых состоит в том, что ауксиллярная клетка, которая формируется незадолго до оплодотворения, является производной несущей клетки карпогонной ветви. Все родимениевые имеют прокарп - ауксиллярная и карпогонная ветви представляют собой единое образование. От несущей клетки развиваются карпогонная ветвь, всегда трех-, четырехклеточная, и двухклеточная ауксиллярная ветвь, верхняя клетка которой служит ауксиллярной. До оплодотворения ауксиллярная клетка плохо различима, но после него сильно увеличивается и наполняется густым плазматическим содержимым. Все водоросли порядка построены по многоосевому типу.
Рост осуществляется в результате деятельности апикальных клеток, а также верхушечной или краевой меристем.
Порядок делится на два семейства, различающиеся особенностями анатомического строения. Семейство шампиевых (Champiaceae) характеризуется полым слоевищем, при этом полость ограничена длинными узкими клеточными нитями. Тетраспорангии делятся тетраэдрически, некоторые роды имеют полиспоры. В семействе родимениевых (Bhodymeniaceae) представители многих родов лишены полости; у тех же, что имеют ее, длинные узкие нити вокруг полости отсутствуют. Тетраспорангии делятся крестообразно.
Несмотря на небольшой объем, порядок отличается чрезвычайно широким ареалом. Родимениевые распространены по всем морям от тропиков до Северного Ледовитого океана, но все-таки наиболее богато они представлены в теплых морях.
Семейство шампиевые (Champiaceae)
Строение водорослей семейства шампиевых удобнее всего разобрать на примере хилокладии чашевидной (Chylocladia kaliformis) - вида, растущего у Атлантического побережья Европы и в Средиземном море. Трубчатое, полое, с перетяжками слоевище хилокладии имеет членистый вид. В местах перетяжек возникают мутовки веточек. Полость, заполненная слизью, прерывается только на перетяжках, так как здесь образуется однослойная клеточная диафрагма. В старых частях слоевища коровой слой состоит преимущественно из одного ряда радиально вытянутых клеток. С внутренней стороны к нему примыкают длинные узкие клеточные нити, спускающиеся вдоль слоевища и расположенные на одинаковом расстоянии друг от друга. Начинаются эти нити у вершины, где расположены апикальные клетки. Каждая апикальная клетка отчленяет вниз сегменты, которые делятся вертикальной перегородкой, образуя две клетки: внешняя формирует коровой слой, внутренняя удлиняется и дает начало продольной нити. Клетки продольных нитей в дальнейшем не подвергаются поперечным делениям, а лишь вытягиваются в длину. Со стороны полости, сбоку от них, отчленяются шаровидные клетки с густым протопластом. Это железистые клетки. Они хорошо заметны в полости слоевища на поперечном срезе не только у хилокладии, но и у других шампиевых. В местах ветвления слоевища железистые клетки делятся, образуя в горизонтальной плоскости клеточные ряды, которые, смыкаясь, складываются в однослойную диафрагму (рис. 180,2).
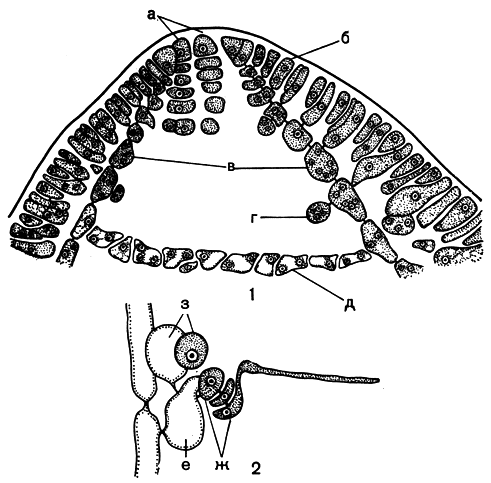
Рис. 180. Строение и размножение шампиевых: 1 - вершина слоевища Chylocladia; 2 - прокарп Lomentaria. а - апикальные клетки, б - клетки коры, в - клетки продольных нитей, г - железистые клетки, д - диафрагма, е - несущая клетка, ж - карпогонная ветвь, з - ауксиллярная ветвь
Другие представители семейства несколько отличаются по внутреннему строению, хотя внешняя перетянутость слоевища в той или иной мере проявляется у всех из них. У шампии (Champia) диафрагма, так же как и кора, может быть многослойной. У видов ломентарии (Lomentaria, табл. 21, 2) диафрагма особого типа. Возникает она за счет того, что вновь образующиеся клетки продольных нитей, вместо того чтобы отклоняться друг от друга в процессе роста, тесно соединяются многочисленными вторичными порами.
Размножение у всех водорослей порядка родимениевых весьма сходно. В семействе шампиевых у ломентарии мелкобулавовидной (L. clavellosa) карпогонная ветвь развивается от коровых клеток, которые соединены с продольными нитями первичными порами. От одной и той же несущей клетки отчленяется трехклеточная карпогонная и двухклеточная ауксиллярная ветви. Верхняя клетка последней является собственно ауксиллярной (рис. 180, 2). Развитие гонимобласта происходит как у родимении. Тетраэдрически разделенные тетраспорангии собраны сорусами в молодых частях растения. Вокруг места их образования коровой слой разрастается, так что группы спорангиев оказываются в небольших углублениях.
Начиная прорастать, споры делятся на четыре клетки. Последующие деления приводят к образованию двухслойного диска. Четыре верхние клетки диска являются первичными апикальными клетками, которые, делясь в горизонтальной плоскости, формируют вертикальный проросток.
Семейство родимениевые (Rhodymeniaceae)
Семейство родимениевых по сравнению с шампиевыми гораздо более многочисленная группа. Ядро семейства составляют тропические и субтропические виды. В морях СССР оно представлено немногими родами, но среди них такие широко распространенные, как родимения и галосакцион.
Род родимения (Bhodymenia) включает около 50 видов, в наших морях их всего четыре. Два близких вида - родимения пальчатая и родимения узкоуголъная (табл. 22, 2) - обычные обитатели литорали умеренных морей. Первый вид растет в наших северных европейских морях, второй - на Дальнем Востоке. Другие два вида встречаются в азиатских морях. Родимения четкообразная не выходит за пределы Охотского моря, тогда как родимения продырявленная (табл. 21, 4) распространена более широко. Это сублиторальный, глубоководный вид.
Пластинчатое слоевище родимении имеет псевдопаренхимное строение. Центральная часть слоевища сложена из нескольких слоев крупных прозрачных клеток, коровой слой плотный, состоит из одного-двух слоев мелких окрашенных клеток. Между корой и сердцевиной имеется небольшая зона клеток промежуточных размеров (рис. 181, 1). У многих более просто устроенных родов семейства коровой слой более рыхлый и состоит из отчетливо различимых клеточных нитей.
Род родимения относится к той группе водорослей семейства, у которых полость внутри слоевища отсутствует, хотя в старых частях слоевища клетки центральной части легко расходятся.
Род галосакцион (Halosaccion) близок к родимении, но если у родимении расхождение клеточных слоев - явление эпизодическое, то у галосакциона наличие полости - постоянный признак, отсутствует она лишь в самых молодых частях слоевища. Однако принцип образо-вания полого слоевища у галосакциона тот же - в результате расхождения клеток в первоначально плотном слоевище. Виды галосакциона - обычные представители литоральной флоры наших морей и часто растут вместе с родименией.
Кроме родов, построенных по типу родимении, в семействе имеется целая группа водорослей с первоначально полым слоевищем. Их строение во многом напоминает строение шампиевых и отличается от них только отсутствием продольных нитей вокруг полости.
У некоторых представителей слоевище может быть просто мешковидно-трубчатым, как, например, у хризимении (табл. 21, 5), у других оно перешнуровано и имеет членистый вид. У ботриокладии полость прослеживается в молодом состоянии и с возрастом заполняется. Полость ограничена слоем крупных клеток, которые переходят в коровой слой, состоящий из нескольких слоев клеток. Вполне возможно, что внутренний крупноклеточный слой формируется в результате того, что клетки продольных нитей, столь характерных для шампиевых, очень сильно увеличиваются и соединяются друг с другом. Это предположение подтверждается тем, что некоторые из внутренних клеток несут железистые клетки (рис. 181, 2).
У родимении несущей клеткой карпогонной ветви служит одна из клеток внутреннего корового слоя, которую можно отличить от соседних по более крупным размерам и богатому содержимому. Карпогонная ветвь трехклеточная. Ауксиллярная клетка формируется до оплодотворения, но только после него становится хорошо различимой, и тогда видно, что это конечная клетка двухклеточной ауксиллярной ветви, сидящей на той же несущей клетке, что и карпогонная ветвь (рис. 181,5). Сразу после оплодотворения в прокарпе происходит ряд изменений. Клетки карпогонной ветви сливаются, затем происходит слияние с ауксиллярной клеткой (рис. 181, 4). Окружающая прокарп ассимиляционная ткань видоизменяется и разрастается, образуя стенки цистокарпа. Большинство клеток гонимобласта при созревании превращаются в карпоспоры, только нижние клетки всегда остаются стерильными. В процессе развития гонимобласта стенки цистокарпа приподнимаются, зрелые цистокарпы имеют полусферическую форму и выступают над поверхностью слоевища. Крестообразные тетраспорангии у родимении разбросаны по слоевищу или собраны в сорусы или нематециевидные образования. Они образуются среди клеток внешней коры и соответствуют одноклеточным боковым веточкам коровых нитей.
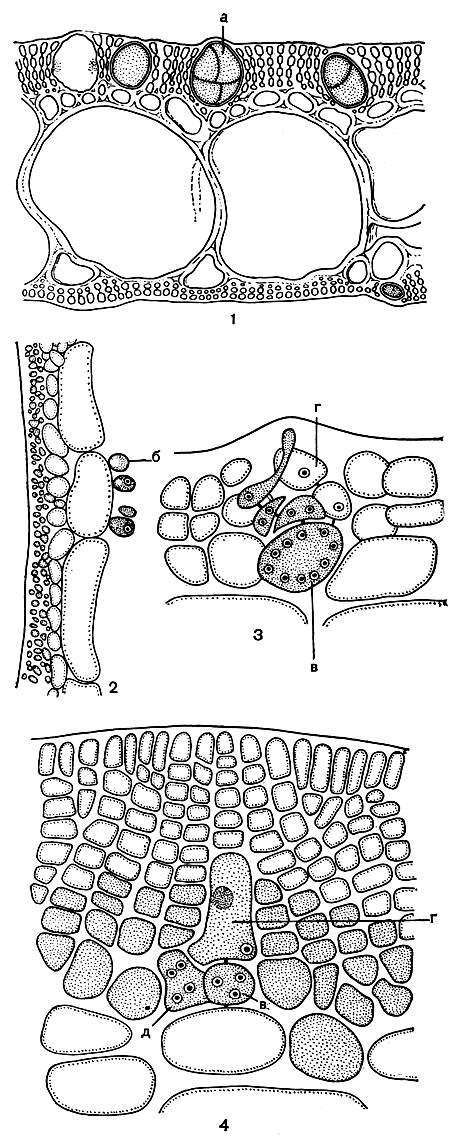
Рис. 181. Строение и размножение родимениевых: 1 - поперечный срез Rhodymenia; 2 - часть среза Chrysymenia; 3 - прокарп Rhodymenia до оплодотворения; 4 - он же после оплодотворения, а - тетраспоры, б - железистые клетки, в - несущая клетка, с - ауксиллярная клетка, д - карпогонная ветвь после слияния ее клеток
Приведенное краткое описание обоих семейств позволяет говорить о большой их близости и рассматривать весь порядок как единую хорошо очерченную группу. Единственное различие между ними заключается в наличии у шампиевых продольных нитей внутри слоевища и в способе деления тетраспорангиев. Судя по тому, что структуру родимениевых можно представить как производную от продольных нитей шампиевых, родимениевые представляют собой более поздний этап эволюции по сравнению с шампиевыми. Однако до конца вопрос о связи обоих семейств, так же как и всего порядка с другими багрянками, остается невыясненным.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'