Порядок криптонемиевые (Cryptonemiales)
У криптонемиевых уже имеются специальные ауксиллярные клетки. Они закладываются до оплодотворения на особых дополнительных ветвях в отдалении от карпогонных ветвей или в непосредственной близости к ним, образуя прокарп. Нити гонимобласта развиваются из ауксиллярных клеток после слияния их с карпогоном.
Слоевище криптонемиевых имеет различную форму (от цилиндрической до пластинчатой и корковидной) и различное анатомическое строение (как одноосевого, так и многоосевого типа). Целая группа водорослей обладает обызвествленным слоевищем. Клетки одноядерные и многоядерные, с одним или несколькими хлоропластами пластинчатой или чечевицеобразной формы, лишенными пиреноидов. Рост осуществляется одной или несколькими апикальными клетками, в формировании слоевища ряда водорослей участвует особая меристема. Для семейства кораллиновых весьма характерен интеркалярный рост.
У криптонемиевых имеется чередование гаметофита и тетраспорофита, сходных по внешнему и внутреннему строению. Тетраспорангии, крестообразно или зонально разделенные, развиваются по-разному: одиночно, в сорусах, нематециях, концептакулах. Сперматангии развиваются так же, как тетраспорангии. Карпогонные ветви простые или разветвленные, обычно многоклеточные; они возникают отдельно друг от друга или по нескольку вместе в сорусах, нематециях или концептакулах.
После церамиевых криптонемиевые - самый крупный порядок красных водорослей. Он включает 13 семейств, 110 родов и более 900 видов, распространенных по всему Мировому океану.
Семейство дюмонтиевые (Dumontiaceae)
Наиболее примитивное семейство дюмонтиевых включает множество родов, встречающихся преимущественно в морях умеренных зон. Они растут в северных частях Атлантического и Тихого океанов, а также у берегов Южной Австралии; гораздо реже встречаются дюмонтиевые в тропической зоне. Более других в отечественных морях известна водоросль дюмонтия (Dumontia, табл. 20, 2). Ее можно встретить на мелководье, часто в литоральных лужах во внутренних частях заливов и бухт. Особенно хорошо развивается дюмонтия в биоценозе двустворчатого моллюска мидии. В скоплениях мидий богатая органика и отсутствие прямого удара волн сочетаются с хорошей аэрацией. Мягкие слизистые растения дюмонтии могут быть длиной 20-60 см. Ее слоевище построено по одноосевому типу (рис. 168, 1). Как и у других водорослей с таким строением, верхушечная клетка поперечной или косой перегородкой отделяет вниз сегменты, от каждого из которых образуется по 4 перицентральных клетки, дающих начало радиально расходящимся пучкам разветвленных клеточных нитей. В верхних частях клетки этих нитей смыкаются и образуется сплошной псевдопаренхимный коровой слой. Единственная центральная ось с возрастом теряется, и слоевище становится полым. Вертикальное слоевище возникает из базальной корки, которая состоит из стелющейся системы однорядных нитей и плотно расположенных вертикальных нитей, как правило, коротких и мало разветвленных. При сильном развитии новые корки могут налегать на старые. В отличие от вертикального слоевища базальная часть многолетняя и служит для переживания неблагоприятных условий.
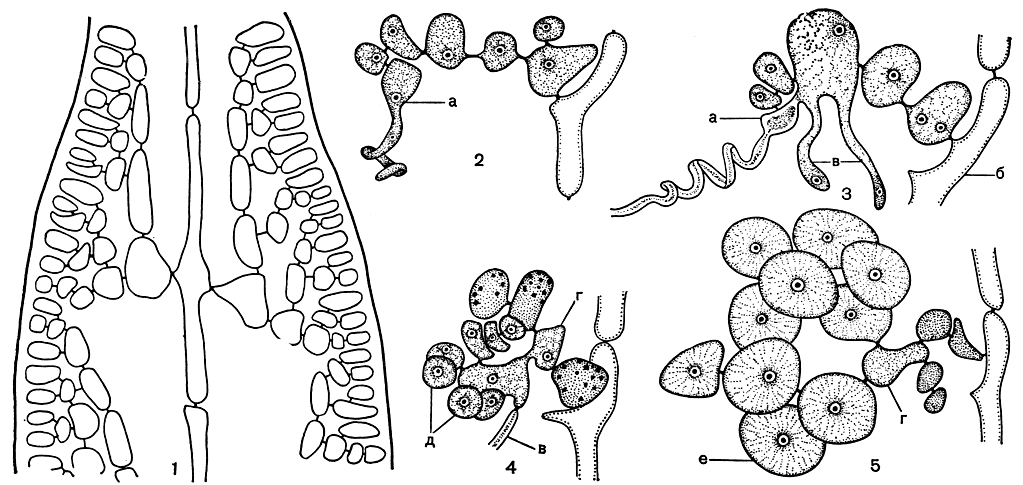
Рис. 168. Строение и размножение Dumontia: 1 - продольный срез у вершины веточки; 2 - карпогонная ветвь; 3 - эта же ветвь после оплодотворения; 4,5 - развитие гонимобласта из ауксиллярной клетки, а - карпогон, б - несущая клетка, в - соединительные нити, г - ауксиллярная клетка, д - первые клетки гонимобласта, е - карпоспоры
При прорастании спор первые деления приводят к образованию шаровидных клеточных структур, которые при дальнейшем развитии превращаются в корковидную подошву. Отдельные нити этой подошвы, разрастаясь, дают вертикальное слоевище дюмонтии.
По сложности организации дюмонтия занимает срединное положение в семействе: в нем можно выделить и роды, построенные более просто, чем дюмонтия, и более специализированные. У низкоорганизованных дюмонтиевых центральная ось покрыта мутовками разветвленных ассимиляционных нитей, которые располагаются свободно и соединены лишь сильно развитой слизью. Эти формы очень напоминают растения батрахоспермума. У более высокоорганизованны видов эти нити смыкаются, образуя сплошной коровой слой. У еще более сложно устроенных дюмонтиевых, как, например, у арктического вида дилсеи съедобной (Dilsea edulis), на ранних стадиях рост пластинчатого слоевища осуществляется верхушечной клеткой, но очень быстро деятельность ее прекращается, а из конечных клеток ассимиляционных нитей, лежащих ближе всего к верхушечной клетке, формируется краевая поверхностная меристема, за счет которой происходит рост растений.
При половом размножении дюмонтии сперматангии образуются на мужских растениях из поверхностных коровых клеток. Материнская клетка сперматангия последовательно отчленяет до трех сперматангиев. При сильном их развитии почти вся поверхность мужских растений бывает покрыта сперматангиями. Обычно мужские растения дюмонтии мельче, чем женские.
Особенности развития женской репродуктивной системы и карпоспорофита дюмонтиевых заключаются в том, что карпогонная и ауксиллярная ветви развиваются по всему слоевищу, во внутренней его части отдельно друг от друга (причем ауксиллярных ветвей больше, чем карпогонных). После оплодотворения карпогон сливается со средней клеткой карпогонной ветви, иногда с несколькими. Из образовавшейся клетки слияния вырастают две или три длинные соединительные нити, которые достигают ауксиллярных ветвей (рис. 168, 2, 3). Собственно ауксиллярной клеткой служит одна из срединных клеток ауксиллярной ветви. После слияния с соединительными нитями из ауксиллярной клетки развивается несколько коротких разветвленных нитей гонимобласта. При созревании все клетки этих нитей превращаются в карпоспоры. Зрелые гонимобласты лишены специальной обертки и погружены в коровую ткань растения (рис. 168, 4, 5). Если проанализировать строение карпогонной и ауксиллярной ветвей у всех дюмонтиевых, становится очевидным, что это структуры гомологичные, т. е. имеющие одинаковое происхождение.
Тетраспорангии образуются из поверхностных клеток коры и разбросаны по слоевищу. В месте образования тетраспорангиев кора толще, чем в стерильной части слоевища. У дюмонтии они крестообразно разделенные, но у других членов семейства встречаются и зонально разделенные тетраспорангии.
Органы полового и бесполого размножения у дюмонтии образуются на сходных по строению макроскопических растениях, т. е. эта водоросль обладает изоморфной сменой форм развития. Это нельзя сказать обо всех членах семейства. У акросимфитона (Acrosymphyton) обнаружен тетраспорофит, который заметно отличается от половых растений и очень напоминает тетраспорофиты некоторых видов боннемезонии.
Дюмонтиевые занимают несколько обособленное положение в порядке криптонемиевых. Они обладают однокарпогонной системой - каждая несущая клетка образует только одну карпогонную ветвь. Все остальные семейства порядка (так полагают в настоящее время) связаны более тесным родством и имеют общего предка, который характеризуется многокарпогонной женской системой, когда на одной несущей клетке развивается несколько карпогонных ветвей. В ходе эволюции эта многокарпогонная система претерпевала изменения и у многих родов вторично преобразовалась в однокарпогонную.
Семейство каллимениевые (Kallymeniaceae)
Семейство каллимениевых является центральным семейством порядка, обладающим многокарпогонным строением женской половой системы. Из шести относимых к семейству родов только три занимают в нем достаточно твердое положение. Это каллимения (Kallymenia), пугетия (Pugetia) и каллофиллис (Callophyllis). Лучше всего изучены виды первого рода, поэтому на них удобнее всего рассмотреть особенности семейства в целом.
Пластинчатые листовидные растения каллимении почковидной (Kallymenia reniformis) можно встретить в северной части Атлантического океана и в Средиземном море. Растет она в сублиторали на значительных глубинах, но доходит и до нижней границы литорали. Молодые растения каллимении состоят обычно из не большой цельной пластины, сидящей на короткой ножке и прикрепленной к грунту дисковидной подошвой. К концу первого сезона роста на пластине развиваются органы размножения. После периода плодоношения большая часть пластины отпадает. В начале следующего сезона по краю оставшейся части пластины возникают пролификации, сама пластина разрывается на лопасти и все слоевище становится многораздельным. Конструкция пластины многоосевая, это можно видеть на поперечном срезе через край пластины (рис. 169, 1). Клетки центральных нитей длинные и узкие. Разветвляясь к поверхности, они образуют внутреннюю кору из одного или двух слоев округлых клеток и внешнюю, также из одного-двух слоев, но уже мелких плотно расположенных клеток. Отдельные клетки центральных нитей очень сильно увеличиваются в размерах и выпускают множество отростков, которые соединяются вторичными порами с обычными клетками и с отростками других таких же клеток. В результате образуется сплошная система звездчатых клеток. Роль этой "ткани", по-видимому, механическая: она помогает довольно тонким пластинам противостоять действию волн. Интересно, что по сравнению с другими пластинчатыми багрянками каллимения чаще сохраняет целостность пластины. Возможно, причина этого кроется как раз в наличии своеобразной механической ткани в центре слоевища. В росте каллимении важную роль играет краевая меристема.
Особый интерес представляет развитие женской половой системы и гонимобласта у каллимении. У нее, как и у многих других водорослей этого порядка, несущая клетка карпогонной ветви возникает от клетки внутренней коры и, увеличиваясь в размерах, ориентируется внутрь слоевища. Каждая несущая клетка отчленяет несколько первичных клеток карпогонных ветвей. Их может быть и шесть, и шестнадцать. Эти клетки, в свою очередь, делятся, образуя трехклеточные карпогонные ветви, и только некоторые из них так и остаются в одноклеточном состоянии, не подвергаясь дальнейшему развитию (рис. 169, 2). Трихогины всех карпогонов одной несущей клетки вытягиваются одновременно. Часто это длинные, спирально закрученные образования. Сразу после оплодотворения происходит кратковременное соединение карпогона с нижней клеткой карпогонной ветви. Вскоре после этого нижние клетки карпогонных ветвей образуют короткие выросты, которые вытягиваются и сливаются с несущей клеткой. Одновременно с этим трихогина, карпогон и подкарпогонная клетка уменьшаются в размерах и постепенно дегенерируют. После соединения нижних клеток всех карпогонных ветвей с несущей образуется очень крупная клетка слияния, которая более чем в 2 раза превышает диаметр несущей клетки. От клетки слияния образуется множество лопастных и трубчатых выростов. Раньше считали, что каллимении свойствен прокарп, так как клетку слияния принимали за ауксиллярную, а лопастные выросты, идущие от клетки слияния,- за первичные нити гонимобласта. Но потом было установлено, что это соединительные нити, растущие в направлении ауксиллярной клетки и сливающиеся с ней. Ауксиллярная система располагается отдельно, независимо от карпогонной. В общей сложности от одной клетки слияния может отходить более десятка соединительных нитей. Они простираются среди нитей центральной части слоевища и могут быть приняты за вегетативные нити. Соединительные нити толще вегетативных, при значительной длине они не разделяются клеточными перегородками, весьма редко ветвятся и не соединяются вторичными порами с вегетативными клетками.
Ауксиллярные клетки также отделяются от клеток внутренней коры. Они очень напоминают несущие клетки карпогонных ветвей. Подобно им, ауксиллярные клетки также отчленяют несколько клеток, которые, однако,дальнейшим делениям не подвергаются. Нет сомнения в том, что ауксиллярная и карпогонная системы - гомологичные образования. Вероятно, первая сформировалась из второй в результате утраты функции оплодотворения и приобретения питающей функции. После слияния с ауксиллярной клеткой соединительная нить вблизи места слияния заметно утолщается и делится, отчленяя от себя первичные клетки гонимобласта. Таким образом, у каллимении гонимобласт берет начало не от ауксиллярной клетки, а от соединительной нити, но только после слияния ее с ауксиллярной клеткой. В зрелом гонимобласте трудно различить нитчатое строение, скорее он имеет вид скопления клеток. По мере созревания гонимобласта окружающая вегетативная ткань раздувается и образуется тонкий перикарп. Клетки гонимобласта превращаются в карпоспоры, крупная ауксиллярная клетка дегенерирует, и остается скопление карпоспор, разбросанных почти по всему слоевищу (рис. 169, 3).
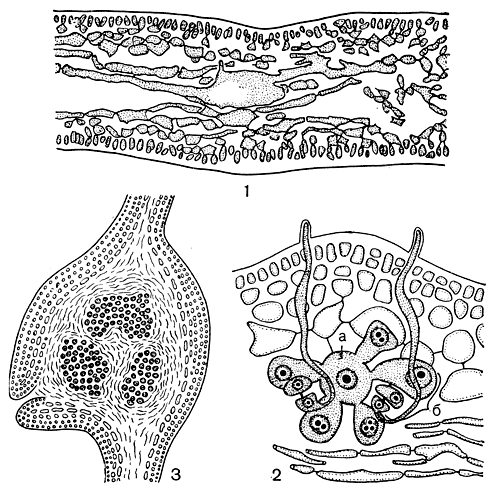
Рис. 169. Kallymenia: 1 - поперечный срез слоевища; 2 - карпогонная система; 3 - поперечный срез через зрелый цистокарп. а - несущая клетка, б - карпогонная ветвь
Крестообразно разделенные тетраспорангии развиваются на бесполых растениях и разбросаны среди клеток внешней коры, хотя развитие их начинается во внутренней коре.
Если сравнить между собой все водоросли семейства, то становится очевидным, что наиболее примитивные члены семейства - это те, у которых внутренняя часть слоевища построена узкими ризоидными нитями, на одной несущей клетке образуется несколько карпогонных ветвей, а карпогонная и ауксиллярная ветви удалены друг от друга. Наоборот, высокоорганизованные представители характеризуются сердцевиной паренхимного вида, одной карпогонной ветвью на несущей клетке и наличием прокарпа. В соответствии с этим каллимению следует считать примитивным представителем семейства, тогда как наиболее специализированным родом оказывается каллофиллис.
Виды каллофиллиса - глубоководные организмы. В нашей стране наибольшего разнообразия они достигают в морях Дальнего Востока, однако общее количество живущих здесь видов назвать весьма затруднительно: обитая в сублиторали труднодоступных районов, этот интересный род остается пока почти неизученным. В переводе на русский язык название этой водоросли звучит как "красиволистный". Плоские пластинчатые слоевища каллофиллиса отличаются от каллимении многократно разветвленной и разделенной пластиной и вследствие этого очень разнообразной формой. Рост слоевища, как у каллимении, происходит в результате деятельности краевой меристемы. На поперечном срезе видно, что внутренняя часть слоевища построена из крупных округлых клеток, между которыми простираются, нередко в большом количестве, тонкие гифообразные клеточные нити (рис. 153, 2).
Развитие карпогонной системы и первые стадии после оплодотворения здесь принципиально не отличаются от этих процессов у каллимении. Но в целом между двумя родами имеются серьезные различия, заключающиеся в том, что несущая клетка у большинства видов дает только одну, а не несколько карпогонных ветвей и гонимобласт начинает развиваться из первой клетки слияния, которая образуется из соединения нижней клетки карпогонной ветви, несущей клетки и дополнительных клеток, отчленившихся ранее от несущей. При таком ходе развития гонимобласта несущая клетка карпогонной ветви является одновременно и ауксиллярной. Иными словами, у каллофиллиса можно говорить о наличии прокарпа.
Мужские растения каллофиллиса мельче женских, так же как и тетраспоровых. Сперматангии образуются по 2-3 от поверхностных клеток внешней коры.
Крестообразно разделенные тетраспорангии разбросаны почти по всему слоевищу. Образуются они от клеток внутренней коры как одноклеточная ветвь.
Семейство криптонемиевые (Cryptonemiaceae)
Водоросли семейства криптонемиевых имеют однокарпогонную половую систему, которая, по-видимому, сформировалась в результате уменьшения числа карпогонных ветвей в первоначально многокарпогонной структуре.
Семейство криптонемиевых - одно из обширных в порядке. В нем известно 19 родов, распространенных в теплых морях. В южные моря Советского Союза заходят лишь очень немногие роды с наиболее широкими ареалами. В первую очередь из них следует упомянуть род грателупия (Grataeloupia). Внешняя форма видов грателупии весьма разнообразна и служит важным критерием для их разграничения. Слоевище кожистое или хрящеватое, крупное, длиной от 5 до 75 см, уплощенное или пластинчатое, сильно и неправильно разветвленное или простое. Построено оно по многоосевому плану. На поперечном срезе через слоевище видно, что внешняя кора состоит из трех-четырех слоев плотно расположенных изодиаметрических клеток. Внутренняя кора более рыхлая, так же как и центральная часть. Клетки центральных осевых нитей имеют длинные отростки своеобразной звездчатой формы. Среди них развиваются многочисленные ризоиды, придающие центральной ткани сетчатое строение (рис. 170, 1).
Женская репродуктивная система располагается на клетках внутренней коры. Несущая клетка двухклеточной карпогонной ветви образует, кроме нее, еще несколько дополнительных клеточных нитей, собранных в виде пучка (рис. 170, 2). Точно так же и в ауксиллярной системе одна базальная клетка несет на себе пучок клеточных нитей, при этом более густой, чем в карпогонной системе. Эта базальная клетка и служит ауксиллярной (рис. 170, 3). Карпогонная и ауксиллярная ветви отделены друг от друга. Прокарп у изученных представителей семейства до сих пор не обнаружен. Хотя отсутствие прокарпа следует считать признаком, свидетельствующим о незначительной специализации, характер карпогонной и ауксиллярной ветвей позволяет думать, что обе происходят из многокарпогонной структуры, которую мы встречаем у каллимениевых. Это предположение вызвано наличием дополнительных клеточных нитей, которые отходят наряду с карпогонной и ауксиллярной ветвями от их несущих клеток. От оплодотворенного карпогона тянутся соединительные нити, которые сливаются с ауксиллярной клеткой (рис. 170, 4). После слияния из нее развиваются разветвленные нити гонимобласта, собранные густым пучком. Созревая, многие клетки гонимобласта превращаются в карпоспоры. Специальной обертки вокруг гонимобласта не образуется, но по мере того, как нити гонимобласта растут к поверхности слоевища, коровая ткань приподнимается и в ней образуется отверстие для выхода карпоспор (рис. 170, 5).
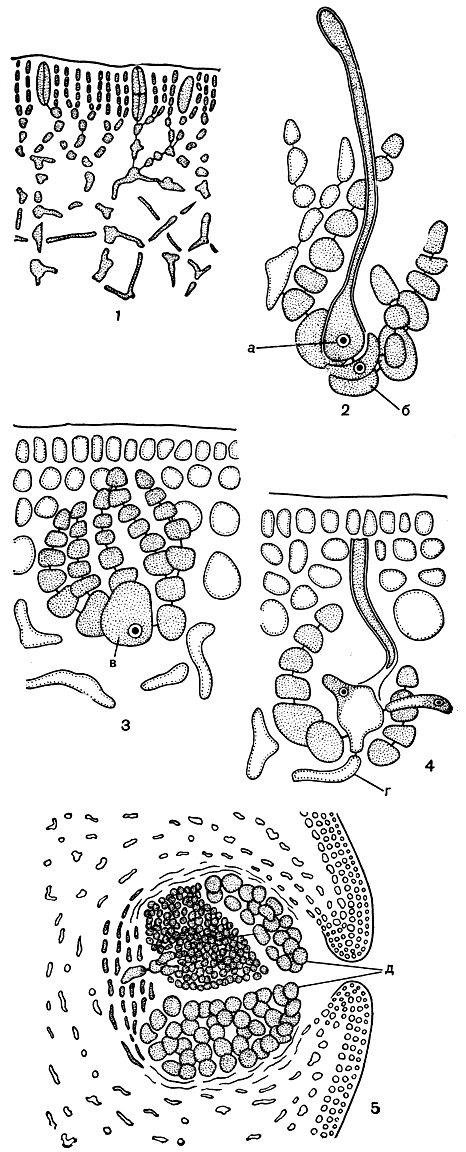
Рис. 170. Строение и размножение Grataeloupia: 1 - поперечный срез слоевища; 2 - карпогонная система; 3 - ауксиллярная система; 4 - оплодотворенный карпогон; 5 - зрелый цистокарп. а - карпогон, б - несущая клетка, в - ауксиллярная клетка, г - соединительные нити, д - карпоспоры
Бесполое размножение осуществляется тетраспорами, которые возникают в крестообразно разделенных спорангиях и разбросаны по всему слоевищу. Иногда в месте образования тетраспор наружная кора утолщается, приобретая нематециевидную форму.
Семейство хореоколаксовые (Choreocolaceae)
Небольшое семейство хореоколаксовых включает два хорошо известных паразитических рода - хореоколакс (Choreocolax) и гарвейелла (Harveiella). Оба рода паразитируют на родомеловых из порядка церамиевых, образуя на их поверхности бесцветные или слабоокрашенные подушечки диаметром до 1 мм. Часть слоевища этих водорослей погружена в ткани хозяина. Она представлена разветвленными нитями, которые соединяются с клетками хозяина вторичными порами или специальными присосками. От эндофитных нитей разрастаются на поверхности подушковидные или бородавчатые образования, устроенные очень просто и состоящие из вильчато-разветвленных нитей, расходящихся веером. К вершине клетки нитей уменьшаются в размерах и, смыкаясь, образуют мелкоклеточную кору.
Систематическое положение семейства хореоколаксовых нельзя считать строго установленным. При развитии женской половой системы несущая клетка, кроме карпогонной ветви, дает начало дополнительным нитям, как у криптонемиевых. Но, с другой стороны, ауксиллярная клетка отделяется от несущей, и только после оплодотворения, а это свойственно церамиевым водорослям. Возможно, это семейство следует исключить из порядка криптонемиевых и поместить в порядок церамиевых, тем более что у красных водорослей паразитические виды всегда оказываются родственными организмам-хозяевам. До тех пор пока вопрос этот не решен, семейство Choreocolaceae по традиции остается в порядке криптонемиевых.
Семейство сквамариевые (Squamariaceae)
Водоросли четырех семейств порядка обладают одним весьма характерным признаком - органы размножения у них развиваются в нематециях, т. е. среди специальных нитчатых образований на поверхности слоевища. Наиболее обширными и своеобразными среди этих семейств являются сквамариевые и кораллиновые.
Семейство сквамариевых характеризуется стелющимся корковидным слоевищем, более или менее плотно прикрепленным к субстрату нижней поверхностью. Корки сквамариевых мягкие, и только очень немногие представители семейства имеют слегка обызвествленное слоевище. У пейссоннелии (Peyssonnelia) вначале образуется однослойная пластина, от клеток которой поднимаются простые или разветвленные вертикальные нити, собранные в плотную псевдопаренхимную ткань. На нижней поверхности слоевища развиваются ризоиды (рис. 171, 1).
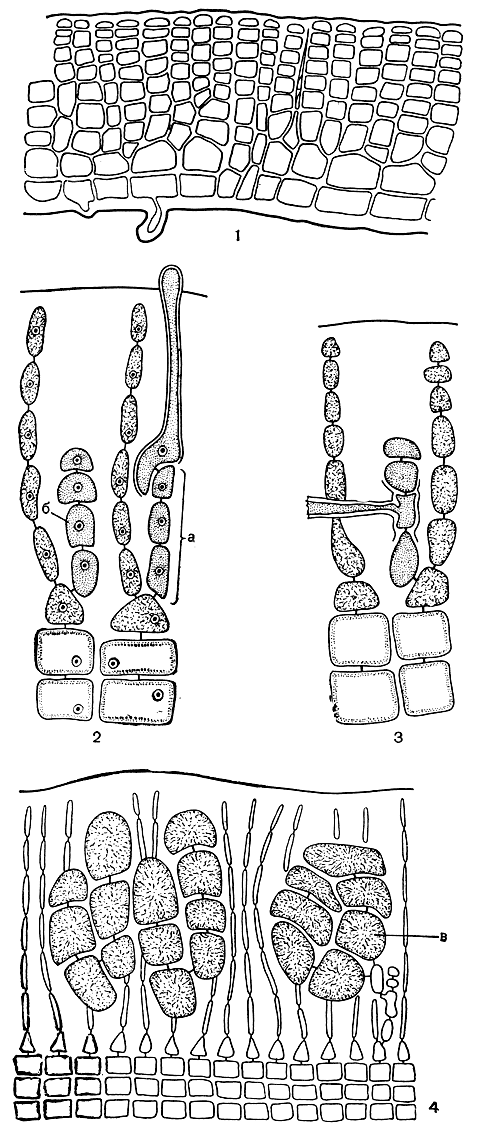
Рис. 171. Строение и размножение Peyssonnelia: 1 - вертикальный срез через корку; 2 - карпогонная (а) и ауксиллярная (б) ветви; 3 - слияние соединительной нити с ауксиллярной клеткой; 4 - гонимобласты. в - карпоспоры
Когда сквамариевые, в частности пейссоннелия, созревают, на поверхности слоевища можно видеть образование нематециев. Группы поверхностных клеток прорастают с образованием многоклеточных вертикальных нитей. При образовании сперматангиев клетки нитей нематеция отчленяют вбок несколько перицентральных, которые служат материнскими клетками сперматангиев (рис. 159, 4).
Нити женских нематециев у пейссоннелии состоят из 6-8 клеток. Самая нижняя клетка такой нити производит карпогонную или ауксиллярную ветвь. Обе ветви, как правило, четырехклеточные (рис. 171, 2). Оплодотворенный карпогон соединяется вначале с двумя клетками карпогонной ветви. Затем эта клетка слияния выпускает соединительные нити, которые соединяются с ауксиллярной клеткой - второй снизу на ауксиллярной ветви (рис. 171, 3). В месте слияния начинается развитие гонимобласта. Он состоит из 8-12 клеток, превращающихся в конечном итоге в карпоспоры (рис. 171, 4).
Крестообразно разделенные тетраспорангии развиваются также среди нитей нематеция от поверхностных клеток слоевища (рис. 158, 5).
Из всех многочисленных водорослей семейства половое размножение известно только у пейссоннелии. Остальные водоросли размножаются тетраспорами. Этот факт позволяет предположить, что сквамариевые, размножающиеся только бесполым путем, являются формами развития других водорослей, а все семейство - искусственной группой. И действительно, в последнее время у некоторых багрянок, в частности у гигартиновых, найдены корковые стадии в циклах развития.
Семейство кораллиновые (Corallinaceae)
Это семейство занимает в порядке особое место. Оно характеризуется огромным многообразием форм. От остальных водорослей порядка, как и вообще от всех багрянок, кораллиновые отличить легко: слоевище их настолько пропитано известью, что их скорее можно принять за кораллы или камни, чем за водоросли. Кроме обызвествления, эти водоросли обладают еще одной своеобразной чертой: органы как полового, так и бесполого размножения у них образуются в особых вместилищах - концептакулах, которые сообщаются с внешней средой одним или несколькими отверстиями.
Систематики делят семейство на два подсемейства, различающиеся морфологией слоевища. Корковые и кораллоподобные водоросли объединяют в подсемейство мелобезиевых (Меlobesioideae), а водоросли с членистым строением слоевища, растущие в форме кустиков, входят в подсемейство кораллиновых (Corallinoideae) (табл. 24). Такое деление водорослей семейства весьма удобно, потому что признак, на котором оно основывается,- внешнее строение - легко бросается в глаза, но вряд ли его можно считать естественным. Различаясь строением взрослого слоевища, водоросли обоих по-семейств обнаруживают сходство по некоторым другим довольно важным признакам. Вероятно, эволюция в семействе шла по нескольким направлениям, каждое из которых включало как корковые, так и членистые формы. Данных об этом сейчас накапливается все больше, но их еще недостаточно, поэтому пересмотр классификации семейства - дело будущего.

Таблица 24. Красные известковые водоросли: вверху - членистые кораллиновые водоросли - кораллина целебная (Corallina officinalis); внизу - корковые кораллиновые водоросли в биоценозе морской травы филлоспадикса. Фото Ю. Астафьева
Как мелобезиевые, так и кораллиновые встречаются во всех морях земного шара от Арктики до тропиков, но более разнообразно представлены в морях теплых широт.
Подсемейство мелобезиевые (Melobesioideae)
Род мелобезия (Melobesia) отличается наиболее простым строением. Виды этого рода растут в виде тонких корочек - чешуек на морских травах, водорослях, раковинах, плотно прилегая к ним всей нижней поверхностью. В начале развития слоевища на субстрате образуется небольшой клеточный диск. Этот диск растет в ширину за счет того, что от его краевых клеток радиально развиваются стелющиеся нити. Они могут располагаться рыхло, но чаще собраны в плотную псевдопаренхимную пластину (рис. 172, 1). Таким образом, на первых порах корка мелобезии состоит из одного слоя клеток. Затем каждая клетка этой однослойной пластины отчленяет наверх по одной мелкой клетке. Эти клетки, если смотреть на пластину сверху, покрывают только небольшой передний участок диска и при этом никогда не обызвествляются. Называют эти клетки покровными. Позднее в клетках диска начинаются деления горизонтальными перегородками и образуются короткие плотно сомкнутые вертикальные нити. Корка становится многослойной (рис. 172, 2). Вертикальные нити растут посредством апикальных клеток и интеркалярно. Мелобезиевые представляют собой один из редких примеров интеркалярного роста у красных водорослей.
Таким образом, в слоевище мелобезии, проследив за его развитием, можно различить два слоя - первичный базальный и вторичный вертикальный. Первый называют - гипоталлием, второй - периталлием. У остальных мелобезиевых принцип строения остается тот же: все они имеют эти два слоя. Не у всех водорослей подсемейства гипоталлий однослойный - количество клеточных слоев в базальной части различно у разных родов и служит важным критерием для их разделения. Когда слоевище сформировано, отличить гипоталлий от периталлия на вертикальном срезе через слоевище можно по форме и цвету клеток, а также по характеру их расположения.
Характерной особенностью мелобезиевых является образование особых клеток, несущих волоски,- трихоцитов. Это крупные, бедные содержимым клетки, лишенные покровных. Они образуются на поверхности слоевища, чаще по краю пластины, но по мере дальнейшего ее роста в ширину обрастают соседними нитями и оказываются во внутренних частях пластины. Волосок не является самостоятельной клеткой, он представляет собой прозрачный более или менее длинный вырост трихоцита (рис. 172, 3). Через некоторое время волосок отпадает, и только перед моментом отпадения в его основании возникает поперечная перегородка. После отпадения волоска трихоциты нетрудно различить по крупным по сравнению с соседними клетками размерам и бедному содержимому.
Клетки гипоталлия, так же как и соседних нитей периталлия, соединяются между собой, но соединение это происходит не посредством вторичных пор, а в результате частичного растворения оболочек и прямого слияния соседних клеток.
С процессом размножения у мелобезиевых связано образование концептакулов. На месте будущего бесполого концептакула из нижних клеток периталлия образуются, чередуясь с обычными вегетативными, фертильные (плодоносные) нити. Каждая такая нить (у мелобезии она может быть очень короткой) оканчивается молодым тетраспорангием, над которым еще в самом начале его образования формируется маленькая клетка с густым содержимым. Вегетативные нити, окружающие спорангии, продолжают расти, но по мере того, как спорангий вытягивается в длину, они дегенерируют и исчезают и таким образом образуется полость концептакула. Нетронутыми остаются только верхние части стерильных нитей, расположенные выше тетраспорангиев. Эта ткань формирует крышу концептакула. Мелкие клетки над тетраспорангиями постепенно теряют цитоплазму и ядро и, оказываясь расположенными среди нитей крыши концептакула, приобретают вид слизистых пробок, закрывающих многочисленные поры, через которые выходят зонально поделенные спорангии, выдавливая при этом пробки (рис. 172, 4).
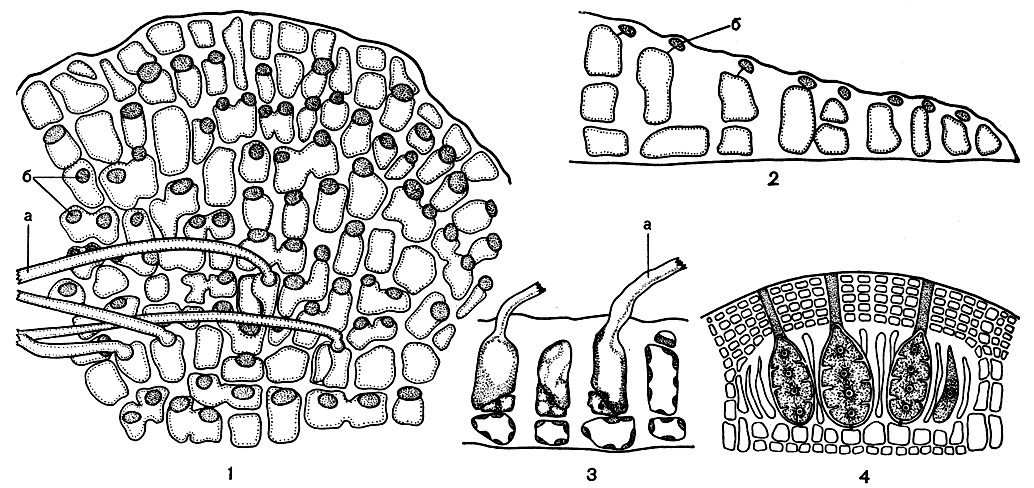
Рис. 172. Строение Melobesia: 1 - корка с поверхности; 2 - вертикальный срез через корку; 3 - трихоциты на вертикальном срезе; 4 - концептакул с тетраспорангиями. а - волоски, б - покровные клетки
Половые концептакулы мелобезии открываются одним отверстием. У мелобезии Лежоли (Melobesia lejolisii) в некоторых местах слоевища клетки, расположенные ниже покровных, начинают делиться с образованием коротких фертильных нитей (рис. 173, 1). В этих нитях от каждой несущей клетки отчленяется наружу по две, очень редко по три мелкие клетки. Одна из них развивается в двухклеточную карпогонную ветвь, другая или другие так и остаются не поделенными. Их считают недоразвитыми карпогонными ветвями. Только изредка можно видеть, как от одной несущей клетки отходит более чем одна карпогонная ветвь. Обычно развитие карпогонных ветвей происходит интенсивнее в центре плодоносного участка (или концептакула), чем на периферии, где обычно карпогонные ветви вообще не развиваются. По мере развития органов размножения окружающие клетки вытягиваются и разрастаются над плодоносным участком в виде клеточной крыши, оставляя лишь одно отверстие в центре. Так формируются женские концептакулы (рис. 173, 2). После оплодотворения карпогон сливается с несущей клеткой, которая таким образом служит ауксиллярной (рис. 173, 3). Затем все ауксиллярные клетки сливаются между собой в одну огромную клетку слияния. Нити гонимобласта вырастают по краю клетки слияния. На конце этих нитей образуются крупные карпоспоры (рис. 173, 4).
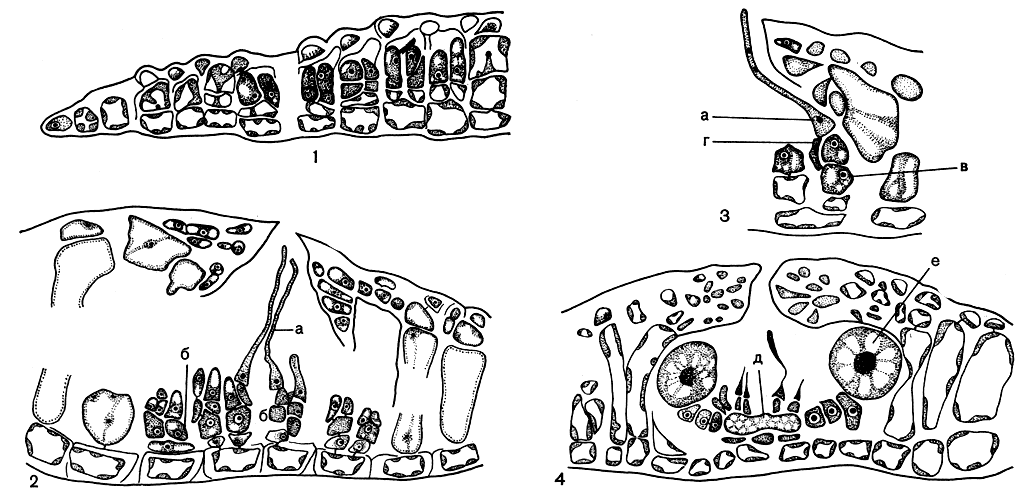
Рис. 173. Развитие гонимобласта Melobesia: 1 - начало образования фертильных нитей; 2 - концептакул со зрелыми карпогонами; 3 - часть концептакула после оплодотворения; 4 - концептакул со зрелым гонимобластом, а - карпогон с трихогиной, б - несущая клетка, в - ауксиллярная клетка, г - соединительная нить, д - клетка слияния, е - карпоспоры
Мужские концептакулы образуются на тех же пластинах, что и женские, в непосредственной близости от них. Из клеток, расположенных ниже покровных, развиваются клеточные нити. Самая нижняя клетка функционирует как клетка-ножка, ряды выше расположенных клеток являются материнскими клетками сперматангиев.
Около 25 видов мелобезии обитает в разных морях земного шара.
Род литотамнион (Lithothamnion) близок к мелобезии. Это название употребляют весьма часто, но не всегда точно. Часто под ним понимают разные известковые водоросли, имеющие вид массивных корок. По сравнению с мелобезией корки литотамниона значительно толще, часто они бородавчатые или снабжены более или менее длинными кораллоподобными выростами. Гипоталлий и периталлий многослойные, покровные клетки крупнее, чем у мелобезии, и покрывают поверхность слоевища целиком. Соединение клеток происходит в результате их слияния. Концептакулы располагаются на поверхности слоевища и устроены по тому же принципу, что у мелобезии.
Виды литотамниона - а их известно около 100 - описаны из различных морей, но многие из этих описаний сомнительны и требуют проверки.
Род литофиллум (Lithophyllum) по внешнему виду не всегда легко отличить от литотамниона. Толщина гипоталлия и периталлия в его многослойном слоевище варьирует таким образом, что отделить этот род от других при отсутствии концептакулов практически невозможно. Несмотря на большое сходство с литотамнионом, между этими родами имеются принципиальные различия. У литофиллума иначе происходит прорастание спор; клетки слоевища соединяются посредством вторичных пор; как бесполые, так и половые концептакулы сообщаются с внешней средой одной порой. Внутреннее строение концептакулов известно не во всех подробностях. В тетраспоровых концептакулах спорангии образуются только по периферии дна концептакула. Карпоспорангии также формируются по краю клетки слияния.
В отличие от литотамниона водоросли из группы литофиллума имеют мелкие споры.
Подсемейство кораллиновые (Corallinoideae)
Членистые кораллиновые характеризуются вертикальным разветвленным слоевищем, в котором обызвествленные членики чередуются с лишенными извести сочленениями. В морях СССР чаще других можно найти кораллину и амфироа. Это наиболее важные представители подсемейства. Каждый из этих родов соединяет в себе признаки целой группы родов и отражает отдельную линию эволюции.
Род кораллина (Corallina) включает невысокие растения, очень красивые благодаря перистому и вильчатому ветвлению (табл. 24; рис. 174,1). Вертикальное слоевище образуется на базальной части, которая здесь развита в меньшей степени, чем у корковых, и намного проще устроена. При прорастании образуется многослойная корка, в которой можно различить гипоталлий и периталлий. Вертикальные побеги построены по фонтанному типу. Членики всегда пропитаны известью, хотя обызвествление обычно не затрагивает внутренних частей и ограничено корой. Нити внутренней части слоевища состоят из клеток одинакового размера, расположенных поперечными рядами (на продольном срезе). Несколько слоев коровых клеток заканчивается у поверхности одним слоем покровных клеток. Последние отсутствуют только над апикальными клетками, осуществляющими рост в длину. Центральные и коровые клетки часто сливаются друг с другом. Сочленение никогда не обызвествляется и состоит из одного ряда параллельно расположенных длинных толстостенных клеток. В начале образования сочленения оно бывает покрыто корой, которая впоследствии разрушается (рис. 174, 2, 3, 4).
У кораллины концептакулы образуются на вершине веточек. При образовании бесполых концептакулов вершины конечных веточек уплощаются, апикальные клетки перестают расти и отчленяют начальные клетки тетраспорангиев. Одновременно с развитием последних происходит разрастание окружающих клеток, в результате которого образуется крыша концептакула. Спорангии делятся зонально (рис. 158, 6). Не все материнские клетки тетраспорангиев достигают зрелости. Часть из них остаются стерильными.
Половые растения кораллины более неправильно разветвлены и гуще покрыты концемтакулами. Развитие женского концептакула начинается с того, что из апикальных клеток центрального пучка нитей образуются плодоносные нити. Их строение и развитие подобны таковым у мелобезии. В зрелом концептакуле различается крупная клетка слияния с нитями гонимобласта по краю. Мужские концептакулы кораллины имеют на вершине довольно длинный клювик, в середине которого проходит длинный выводной канал. Сперматангии образуются на дне и боковых стенках концептакула и имеют весьма характерную форму. Они расширены (наподобие головки) на переднем конце и вытянуты в длинный узкий хвост на заднем.
Род амфироа (Amphiroa) отличается меньшим числом видов и более ограниченным географическим распространением. Вильчато-разветвленные растения амфироа также состоят из отчетливых члеников, однако в их образовании прослеживаются еще некоторые примитивные черты, сближающие этот род с нечленистыми кораллиновыми. Клетки центральных нитей расположены упорядоченно, дугообразными поперечными зонами, причем зона длинных клеток чередуется с зоной коротких. Снаружи центральный пучок ограничен мелкоклеточной корой. Но если у кораллины первое сочленение формируется очень рано, одновременно с возникновением вертикального слоевища, то у амфироа сочленения возникают, когда ткани проростков уже достаточно сильно дифференцировались. Сочленения состоят из нескольких рядов клеток. Имеются вторичные поры (рис. 174, 5).
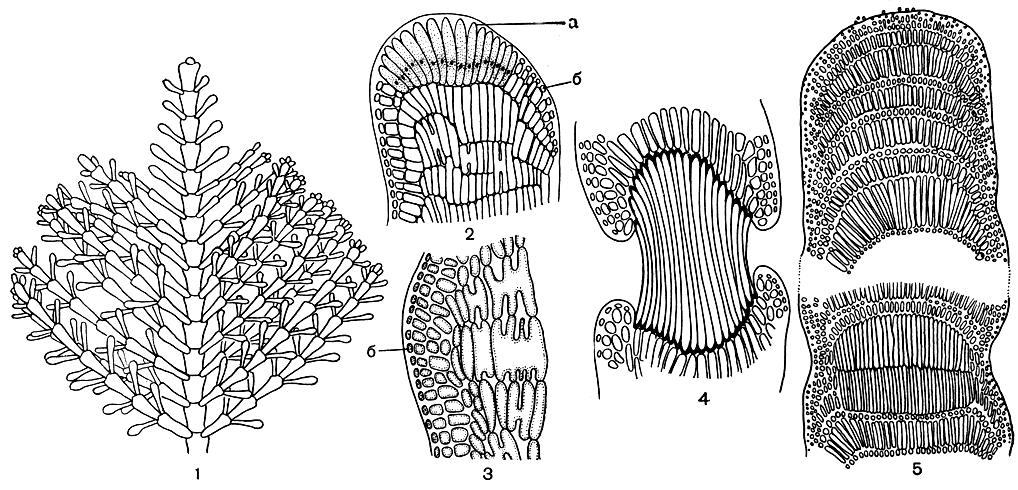
Рис. 174. Строение кораллиновых: 1 - внешний вид веточки Corallina; 2-5 - продольные срезы (2 - через вершину веточки Corallina, 3 - через членик, 4 - через сочленение, 5 - через вершину веточки Amphiroa - видны зоны коротких и длинных клеток и двухрядное сочленение). а - апикальные клетки, б - покровные клетки
Концептакулы располагаются на боковой поверхности ветвей и формируются за счет коровых клеток. На начало развития любого концептакула указывает секреция поверхностными клетками слизистых веществ, которые образуют своего рода шапку в месте будущего концептакула. В то же самое время начинает формироваться свод концептакула. Группа коровых клеток, лежащих под поверхностными, вытягивается перпендикулярно поверхности слоевища. Это клетки, за счет которых позднее образуется полость концептакула. Над ними продолжается образование мелких коровых клеток, формирующих свод концептакула. Дальнейшее развитие бесполых и половых концептакулов идет разными путями. Тетраспорангии берут начало от коровых клеток, лежащих на дне концептакула, и располагаются кольцом по периферии дна концептакула. По мере созревания тетраспорангиев длинные клетки над ними разрушаются с образованием полости. В процессе развития мужских концептакулов с момента возникновения материнских клеток сперматангиев начинается разрушение будущей полости концептакула, так же как и клеток первичного свода, так что крыша сформированного концептакула образуется вторично в результате разрастания окружающих тканей. Сперматангии - мелкие овальные клетки - развиваются на дне концептакула. Женские концептакулы формируются так же, как мужские. В зрелом концептакуле можно видеть крупную клетку слияния с кольцом нитей гонимобласта, отходящих по ее краю.
Стараясь понять филогенез криптонемиевых, ученые исходят из того, что различные направления развития в порядке (впрочем, как и во всем классе флоридеевых) находят отражение прежде всего в характере строения женской половой системы и в особенностях развития гонимобласта. Основными критериями служат наличие или отсутствие прокарпа, число клеток в карпогонной и ауксиллярной ветвях, место образования ауксиллярной клетки.
Раньше все семейства порядка делили на две группы - имеющие прокарп и не имеющие такового. Но чем глубже изучают эти водоросли, тем очевиднее становится, что специализация ауксиллярной системы может идти у разных водорослей независимо. И действительно, во многих семействах наряду с родами, у которых карпогонная и ауксиллярная ветви образуются отдельно, можно найти роды, уже имеющие в той или иной мере развитый прокарп. По современным воззрениям (заметим, что их также нельзя считать непогрешимыми), одним из важных признаков, на который опираются при построении классификации криптонемиевых, является одно- или многокарпогонный характер женской половой системы. Считается, что многокарпогонный аппарат, свойственный многим криптонемиевым, возник очень давно из однокарпогонного, но позднее у некоторых родов однокарпогонная система могла возникнуть вновь в результате редукции многокарпогонной. Наиболее примитивным семейством порядка считают дюмонтиевые. Они имеют ауксиллярные ветви, изолированные от карпогонных, и оба типа ветвей состоят из большого числа клеток. Однокарпогонная система здесь - первичное образование. Наиболее отчетливо многокарпогонная структура проявляется в семействе каллимениевых; в других семействах с одной карпогонной ветвью на несущей клетке наличие на ней же нескольких стерильных клеточных нитей свидетельствует о том, что, по крайней мере в некоторых случаях, эта структура образовалась в результате редукции числа карпогонных ветвей.
Таким образом, эволюция в пределах порядка криптонемиевых шла в направлении образования, во-первых, прокарпа, во-вторых, многокарпогонной структуры, а затем вторичного ее упрощения и, в-третьих, усложнения анатомического строения. Как известно, эволюция различных признаков происходит независимо, и в порядке криптонемиевых сплошь и рядом можно встретить водоросли, которые по одним признакам следует считать высокоорганизованными, тогда как по другим - примитивными. В результате оказывается, что понять эволюцию группы в целом и истинные связи семейств друг с другом трудно. Несомненно, современ-ную классификацию порядка нельзя признать до конца разработанной.
Связь порядка с другими багрянками также остается неясной, хотя имеются свидетельства сходства отдельных криптонемиевых с водорослями порядка гигартиновых.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'