Порядок хордариевые (Chordariales)
К хордариевым относятся бурые водоросли, имеющие самые разнообразные микроскопические и макроскопические слоевища: корковидные, пузыревидные, шнуровидные, кустистые (высотой до 70 см). Общим признаком для хордариевых служит строение поверхностного слоя слоевища. Он состоит из многоклеточных (2-12 клеток) однорядных неразветвленных нитей. Их клетки, содержащие большое количество хлоропластов, интенсивно окрашены; такие нити называют ассимиляционными. Нередко они имеют булавовидную форму за счет того, что их верхушечные клетки крупнее нижних. Ассимиляционные нити бывают рыхло- или плотно расположенными (рис. 125, 127, 2), но они не срастаются между собой. У некоторых представителей, например у стилофоры, ассимиляционные нити появляются группами только вместе с органами размножения; у других ассимиляционные нити выражены только на растущих вершинах (у сплахнидиума).
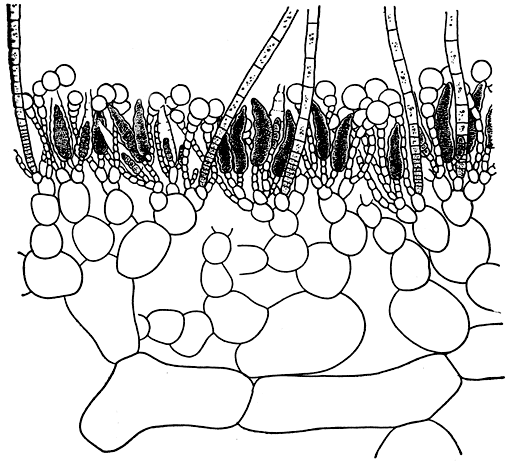
Рис. 125. Продольный разрез слоевища мезоглойи (Mesogloia)
Слоевища хордариевых построены следующим образом. В основании имеется однослойная или многослойная пластинка или разветвленные стелющиеся нити. От основания вверх отходит большое число однорядных многоклеточных вертикальных нитей (при многоосевом строении) или одна нить (при одноосевом строении). Вертикальные нити дают короткие боковые выросты, которые многократно и интенсивно ветвятся, образуя слои клеток, окружающих центральный пучок вертикальных нитей или единственную центральную нить. Различия в строении слоевищ хордариевых связаны с разным числом вертикальных нитей и их высотой, а также с неодинаковой плотностью их расположения. Если вертикальные нити не развиваются, получается корковое слоевище или слоевище, образованное стелющимися нитями. Слоевища большинства хордариевых несут волоски с базальной зоной роста. У некоторых мелких представителей (семейство элахистовых) наряду с ассимиляционными нитями растут периферические нити. Они тоже однорядные, но гораздо длиннее ассимиляционных (до 2,5 см), и их клетки также содержат много хлоропластов.
Цикл развития хордариевых заключается в чередовании большей частью крупного спорофита и микроскопического гаметоспорофита. У хордариевых имеется плетизмоталлюс - микроскопический спорофит, который несет многогнездные спорангии и может в виде выроста производить макроскопическое слоевище спорофита. Из зооспор снова вырастают слоевища плетизмоталлюса. Макроскопические слоевища спорофитов у хордариевых производят одногнездные или многогнездные спорангии или и те и другие одновременно. Мейоз происходит в одногнездных спорангиях. Гаметоспорофит хордариевых состоит их стелющихся разветвленных нитей, на которых развиваются многогнездные вместилища. Половой процесс - изогамия, крайне редко - гетерогамия. Зигота прорастает в растеньице, которое сначала внешне подобно гаметоспорофиту, а позднее производит макроскопическое слоевище спорофита или остается в виде плетизмоталлюса. Зооиды без полового процесса снова дают начало гаметоспорофитам. Из спор, производимых макроскопическими спорофитами, могут вырастать как гаметоспорофиты, так и спорофиты. Все это приводит к тому, что в цикле развития хордариевых отсутствует строгое чередование форм развития.
Из-за большого разнообразия в строении слоевищ хордариевые делят на несколько семейств.
Семейство мирионемовые (Myrionemataceae)
Слоевище у мирионемовых мелкое, микроскопическое, состоящее из одно- или двухслойной стелющейся пластинки с короткими вертикальными нитями (рис. 121, 2). Представители этого семейства часто встречаются на крупных водорослях, где их можно обнаружить только с помощью микроскопа, а также на камнях. Мирионемовые произрастают в литоральной и сублиторальной зонах всех морей. Возможно, что часть водорослей, относящихся к этому семейству, представляют собой микроскопические стадии развития других бурых водорослей.
Особенно часто встречаются виды трех родов.
Род мирионема (Myrionema) включает виды с дисковидным или полусферическим слоевищем, достигающим в диаметре 0,5-4 мм. Основание однослойное, из радиально расходящихся плотно сомкнутых нитей. Высота вертикальных нитей достигает 0,2 мм. Спорофит и гаметоспорофит могут иметь вертикальные нити разного облика и размера. Гаметоспорофиты с многогнездными многорядными вместилищами, спорофиты с одногнездными эллипсоидными или грушевидными спорангиями (рис. 121, 2).
Род аскоциклус (Ascocyclus) отличается от рода мирионема тем, что у его представителей на слоевищах развиваются одноклеточные парафизы. Спорангии известны только однорядные многогнездные.
Род микроспонгиум (Microspongium) объединяет водоросли, у которых слоевища образуют выпуклые слизистые подушечки. В отличие от вышеперечисленных родов вместилища образуются не на поверхности горизонтальной части, а по бокам вертикальных нитей.
Семейство ральфсиевые (Ralfsiaceae)
У ральфсиевых слоевище корковидное, грубое, большей частью многолетнее. Оно состоит из горизонтального слоя клеток, образованного радиально расходящимися нитями, и из вертикальных плотно сомкнутых рядов клеток (рис. 127,2). Одногнездные спорангии и многогнездные вместилища образуются как выросты поверхностных клеток. Одновременно с органами размножения развиваются многоклеточные парафизы. Ральфсиевые растут в морях умеренных и приполярных поясов. Наиболее широко известна ралъфсия бородавчатая (Ralfsia verrucosa). Ее темно-коричневые корки достигают в диаметре 10 см и в толщину 2 мм, на них часто выражены концентрические полосы, соответствующие, видимо, периодам интенсивного роста. Ральфсия бородавчатая растет в литоральной зоне в "ваннах" (углубления в скалах, заполненные водой) и на открытых местах, а также в верхней сублиторали. В бесприливных морях она покрывает камни, омываемые прибоем. Органы размножения наблюдаются летом и осенью.
Семейство литодермовые (Lithodermataceae)
Литодермовые подобны ральфсиевым, но отличаются от них отсутствием парафиз и особенностями развития органов размножения. Одногнездные спорангии образуются из поверхностных клеток, а многогнездные вместилища появляются или как боковые выросты вертикальных нитей, которые вверху обычно неплотно сомкнуты, или путем преобразования их верхних клеток. Среди литодермовых наряду с морскими видами имеются представители, растущие в сильно опресненных местах. Виды рода херибаудиелла (Heribaudiella) встречаются в реках в местах с сильным течением.
Семейство элахистовые (Elachistaceae)
Слоевище элахистовых имеет вид маленького пушистого или плотного шарика, несущего густой пучок нитей. Внутри слоевище состоит из вильчато-ветвящихся коротких нитей, которые у поверхности переходят в неразветвленные короткие ассимиляционные и длинные периферические нити; последние при сильном развитии придают слоевищу вид пучка нитей. У некоторых представителей ассимиляционные нити отсутствуют. Органы размножения развиваются на основных нитях рядом с ассимиляционными или на периферических нитях. Большинство элахистовых растет в качестве эпифитов на крупных водорослях, некоторые имеют снизу бесцветные нити, внедряющиеся в ткань хозяина. Распространены элахистовые в холодных и умеренных морях.
Род элахиста (Elachistea) и его вид элахиста фукусовая (Е. fucicola) наиболее широко известны. Для этих водорослей характерны плотная полушаровидная часть в основании и длинные периферические нити, достигающие в длину 1,5-2 см. Периферические нити появляются летом, а осенью опадают, поэтому зимой и весной слоевище может иметь вид плотных шариков. Органами размножения являются одногнездные и многогнездные (однорядные и многорядные) спорангии. Зооспоры из многогнездных спорангиев служат для воспроизведения материнских растений. В одногнездных спорангиях происходит мейоз; образующиеся зооспоры дают микроскопические гаметоспорофиты, имеющие вид стелющихся разветвленных нитей; на них развиваются многогнездные гаметангии. Переход от аметоспорофитов к спорофитам осуществляется в результате полового процесса и образования зигот или путем развития слоевища спорофита из выроста на гаметоспорофите.
Элахиста фукусовая встречается на бурых водорослях - фукусе и аскофиллуме, в защищенных и открытых местах в литорали и в верхней сублиторали. Особенно в больших количествах она развивается на видах рода фукус, благодаря чему и получила такое видовое название.
Элахиста фукусовая поселяется и на слоевищах красных водорослей - халосакционе и родимении, растущих в тех же местах. Распространена она в северной части Атлантического океана и в арктических морях.
Семейство коринофлеевые (Corynophlaeaceae)
У коринофлеевых слоевище шаровидное, внутренняя часть его образована дихотомически-ветвящимися нитями, построенными из крупных бесцветных клеток. Снаружи располагаются ассимиляционные нити, а периферические нити, в отличие от элахистовых, отсутствуют. Наиболее крупным слоевищем обладает леатезия неоднородная (Leathesia difformis, рис. 129, 2). Она может достигать в диаметре 5 см. В крупных слоевищах внутренние нити разрушаются, и слоевища превращаются в полые шары. На одном слоевище могут одновременно развиваться одногнездные и однорядные многогнездные спорангии. Растет леатезия на камнях и на водорослях в литорали и в верхней сублиторали морей умеренного пояса.
Семейство хордариевые (Chordariaceae)
Слоевище у хордариевых крупное, высотой до 70 см, кустистое, с цилиндрическими ветвями. В центральной части побегов располагается пучок продольных нитей или одна нить. Поверхность слоевища образована ассимиляционными нитями. В отдельных районах морей СССР широко распространены виды пяти родов.
Род эудесме (Eudesme) характеризуется нежным слоевищем с побегами в виде слабо разветвленных слизистых шнуров рыхлого строения (в препарате при надавливании на покровное стекло нити легко расходятся). Ассимиляционные нити располагаются пучками. Центральная часть ветвей сложена пучком нитей, слабо соединенных между собой. Органы размножения - одногнездные спорангии.
Из видов этого рода особенно известен эудесме зеленоватый (Е. virescens). В морях СССР его макроскопические слоевища, являющиеся спорофитами, растут в наиболее теплые летние месяцы в защищенных местах на других водорослях и камнях. Зимой он существует в виде микроскопических гаметоспорофитов, на которых образуются одногнездные спорангии.
Род мезоглойя (Mesogloia) отличается тем, что у его представителей в центре ветвей расположена одна нить, а не пучок нитей (рис. 125). Эти водоросли также имеют рыхлое строение. У спорофитов некоторых видов, кроме одногнездных спорангиев, встречаются многогнездные. Образующиеся в них зооспоры снова дают материнские растения.
Род хордария (Chordaria) включает виды с кустистым слоевищем плотного строения, клеточные нити внутри его (кроме ассимиляционных) плотно соединены между собой (рис. 126, 127, 1). Центральная часть побегов образована пучком нитей обычного для хордариевых диаметра; между ними расположены узкие нити из длинных клеток с толстыми оболочками - гифы, они повышают прочность слоевища на разрыв. Органы размножения - одногнездные спорангии.

Рис. 126. Хордария плетевидная (Chordaria flagelliformis)
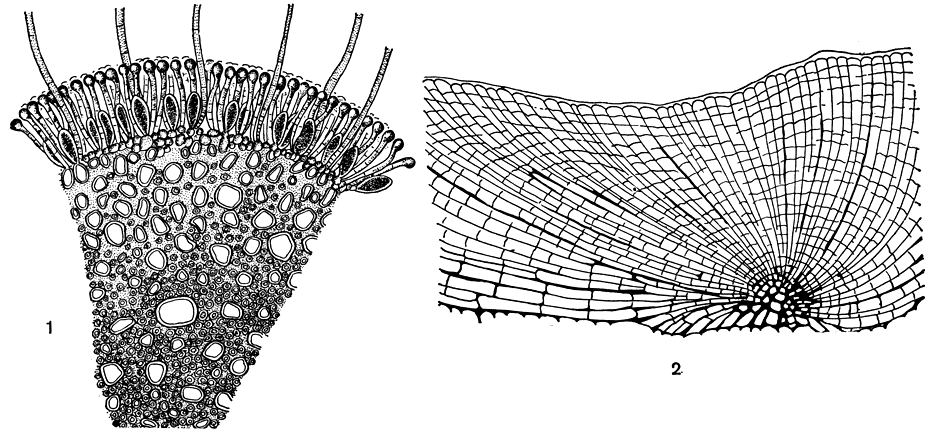
Рис. 127. Поперечный разрез слоевища хордарии плетевидной (1); вертикальный разрез слоевища ральфсии бородавчатой (2)
Особенно широкое распространение имеет хордария плетевидная (С. flagelliformis). Она растет в северном полушарии во всех морях субарктического и умеренного поясов, нередко ее заросли покрывают большие участки каменистого грунта в открытых и защищенных местах. В зависимости от силы прибоя слоевища хордарии плетевидной могут быть то сильно разветвленными (в более спокойных местах), то почти неразветвленными (там, где прибой сильный). Спорангии у спорофитов имеются с апреля до ноября. Гаметоспорофиты микроскопические с многогнездными вместилищами. В условиях лабораторных культур из зооспор, которые производят спорофиты, снова вырастали спорофиты, а также гаметоспорофиты. Последние в культуре продолжали размножаться бесполым путем посредством зооспор. Замечено, что органы размножения на гаметоспорофитах развиваются при более низкой температуре, чем на спорофитах, что соответствует их произрастанию в природе в зимнее время.
Род сферотрихия (Sphaerotrichia) отчасти сходен с предыдущим. Виды этих двух родов легко спутать по внешнему облику, но они хорошо различаются по анатомическому строению. У сферотрихии развесистой (S. divaricata), которая распространена там же, где и хордария плетевидная, отсутствуют гифы. По этому она растет в более спокойных местах, и ее слоевища менее грубые.
Род аналипус (Analipus) удобно рассмотреть на примере наиболее известного вида аналипуса японского (A. japonicus). Он распространен в северной части Тихого океана, в морях умеренного пояса и до недавнего времени больше был известен как гетерохордария еловая (Неterochordaria abietina). В СССР это довольно обычное растение в некоторых районах Японского моря, у Сахалина, Курильских островов и восточного побережья Камчатки. Аналипус японский имеет вертикальное слоевище высотой до 30 см с центральным побегом, от которого отходят короткие неветвящиеся веточки. Коровой слой образован ассимиляционными нитями. Многогнездные вместилища развиваются путем преобразования клеток ассимиляционных нитей, кроме верхушечной клетки. Вертикальное слоевище прикрепляется к грунту подошвой. Иногда оно отходит от стелющихся разветвленных многорядных побегов с плотным коровым слоем. В местах с сильным прибоем вертикальные побеги образуют небольшое число коротких боковых побегов. Могут вырастать вертикальные побеги совершенно без боковых ветвей. В еще более прибойных местах развиваются только стелющиеся побеги или слоевища в виде корок, как у ральфсии.
Семейство сперматохновые (Spermatochnaceae)
Слоевище у сперматохновых кустистое, с одной или четырьмя-пятью осевыми нитями. Коровой слой плотный. Ассимиляционные нити образуются только при развитии органов размножения и выполняют функцию парафиз - бесплодных клеточных нитей. Из этого семейства наиболее широко известны род сперматохнус (Spermatochnus) с одной осевой нитью и род стилофора (Stilophora) с четырьмя-пятью осевыми нитями. В СССР виды этих родов произрастают в сублиторали Черного моря.
Семейство сплахнидиевые (Splachnidiaceae)
К этому семейству относится всего один род сплахнидиум (Splachnidium) с единственным видом. Сплахнидиум морщинистый (Splachnidium rugosum) растет в южном полушарии у мыса Доброй Надежды и по южному берегу Австралии. Слоевище у этого растения студенистое, с полостью внутри; оно состоит из центрального толстого вертикального побега с немногочисленными слабо разветвленными толстыми боковыми ветвями. В основании слоевища имеется диск для прикрепления к грунту. Органы размножения - одногнездные спорангии - у сплахнидиума морщинистого развиваются в особых углублениях - концептакулах. Концептакулы образуются в некоторых точках поверхности слоевища, вблизи вершин побегов. Здесь начинаются интенсивное деление и рост клеток, при этом возникают впячивания внутрь слоевища. Образованию концептакулов способствует то, что слоевище имеет рыхлое нитчатое строение и снабжено полостями, заполненными слизью. В прошлом сплахнидиум морщинистый относили к фукусовым на том основании, что у него существуют концептакулы и на вершинах побегов расположены особые крупные клетки. Однако со временем выяснилось, что концептакулы у них образуются совершенно иначе, чем у фукусовых, а особые клетки оказались одноклеточной эндофитной зеленой водорослью кодиолум (Codiolum), которая постоянно присутствует в коровом слое молодых вершин побегов. Ассимиляционные нити у сплахнидиума морщинистого развиваются только на молодых частях, потом они опадают, и поверхность оказывается образованной плотной корой из мелких клеток. У сплахнидиума имеются многоклеточные волоски с интеркалярной зоной роста, которые растут на внутренней поверхности концептакулов и высовываются через их отверстия наружу.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'