Порядок эктокарповые (Ectocarpales)
К эктокарповым относятся бурые водоросли, слоевища которых построены из однорядных нитей. Эти нити могут ветвиться и образовывать кусты высотой до 30, реже до 60 см, или развиваться в виде микроскопического налета на скалах и других водорослях, или расти в теле крупных водорослей и только в период размножения выходить на поверхность и образовывать органы размножения. Клетки вегетативных нитей эктокарповых имеют окрашенные хлоропласты, содержащие пиреноиды. Органами размножения у этих водорослей служат одногнездные спорангии и многогнездные вместилища, последние могут быть спорангиями или гаметангиями. Половой процесс изо- или гетерогамный. Гетерогамия наблюдается у видов рода гиффордия (Giffordia); у них известно три типа многогнездных вместилищ, различающихся размерами гнезд (микро-, мезо- и мегавместилища). Вместилища с самыми мелкими и с самыми крупными гнездами служат гаметангиями, с гнездами промежуточного размера - спорангиями. У эктокарповых наблюдается вегетативное размножение обрывками нитей.
Эктокарповые встречаются во всех морях мира, но особенно много их в холодных морях и в субтропических морях в холодное время года. Эктокарповые обитают в литорали и в сублиторали, на твердых и мягких грунтах и на более крупных водорослях. Они чаще других бурых водорослей участвуют в обрастании морских судов. В порядок входит только одно семейство эктокарповых (Ectocarpaceae), объединяющее несколько родов.
Род эктокарпус (Ectocarpus) имеет кустистые слоевища высотой 0,1-30 см, крайне редко до 60 см, образованные тонкими нитями толщиной 50-150 мкм. Основанием служат стелющиеся ризоиды; у некоторых видов, растущих на водорослях, они могут проникать внутрь их. Крупные вертикальные побеги в основании бывают покрыты корой из ризоидов. Ветви слоевища сужаются к концам и на вершине оканчиваются длинными бесцветными клетками. Волоски со вставочной зоной роста отсутствуют. Вертикальные побеги растут за счет деления большого числа клеток, особых зон роста нет. У нитей ризоидов существует верхушечный рост. Клетки эктокарпуса снабжены пластинчатыми хлоропластами, в том числе лентовидными (рис. 122, 3). Спорангии и гаметангии располагаются как боковые выросты ветвей, многогнездные вместилища иногда имеют волосовидную стерильную вершину. Цикл развития у видов рода эктокарпус может быть изоморфным или гетероморфным. При этом виды, считающиеся ныне самостоятельными, иногда оказываются формами развития одного вида. Половой процесс всегда изогамный.
Спорофиты эктокарпуса пучковатого (Е. fasciculatus) в Атлантическом океане у северных берегов Франции при + 13 °С несут только одногнездные спорангии, при +20 °С образуются многогнездные спорангии и в небольшом количестве одногнездные. Споры из многогнездных спорангиев снова дают спорофиты. В одногнездных спорангиях происходит мейоз, и из спор вырастают раздельнополые гаметофиты, напоминающие эктокарпус драпарналъдиевидный (Е. draparnaldioides). Оплодотворение происходит после прикрепления женских гамет. Из зигот вырастают спорофиты. При партеногенетическом прорастании гамет (без оплодотворения) появляются растения, подобные спорофитам или гаметофитам.
Образование разных органов размножения и изменение цикла развития в зависимости от района произрастания наблюдаются у эктокарпуса стручковатого (Е. siliculosus, рис. 122, 2). В Белом море и у берегов Норвегии встречаются почти исключительно растения с многогнездными спорангиями - это спорофиты. Около Неаполя зимой растут диплоидные растения с одногнездными спорангиями, производящими в результате мейоза гаплоидные зооспоры. Весной существуют гаплоидные растения с многогнездными вместилищами; образующиеся в них зооиды функционируют как гаметы или как зооспоры. Летом эти растения сменяются диплоидными растениями с одногнездными и многогнездными спорангиями. Из зооспор многогнездных спорангиев вырастают слоевища, растущие зимой и несущие одногнездные спорангии. Из зооспор одногнездных спорангиев вырастают гаметофиты, которых сменяют растущие зимой диплоидные спорофиты. У эктокарпуса стручковатого из Неаполя при культивировании в лаборатории наблюдались гаплоидные и диплоидные неветвящиеся спорофиты, кроме того, были обнаружены диплоидные гаметофиты и тетраплоидные спорофиты. У берегов Новой Англии (Атлантическое побережье США) эктокарпус стручковатый весной представлен диплоидными растениями с одногнездными и многогнездными спорангиями, которые сменяются летом гаметофитами, а осенью и зимой растут диплоидные спорофиты с многогнездными спорангиями. Зооспоры эктокарпуса стручковатого из одногнездных и многогнездных спорангиев различаются строением, однако глазок и жгутики у них одинаковые. Зооспоры из одногнездных спорангиев крупнее, плавают медленнее и реже меняют направление движения.
Род пилайелла (Pylaiella) напоминает эктокарпус (рис. 122, 1). Клетки с мелкими дисковидными хлоропластами. Ветвление одностороннее и супротивное. Одногнездные спорангии образуются цепочками из вегетативных клеток ветвей. В пределах одной цепочки спорангии созревают одновременно; содержимое их выходит последовательно, начиная с верхнего спорангия, через боковые отверстия, открывающиеся на одну сторону, в каждом спорангии по одному отверстию. Многогнездные спорангии многоклеточные, тоже вставочные или в виде боковых веточек. У этих водорослей изредка происходят продольные деления вегетативных клеток, поэтому пилайеллу некоторые исследователи относят к порядку диктиосифоновых. Цикл развития у пилайеллы изоморфный.
Наиболее широко известна пилайелла прибрежная (P. litoralis). У этого вида растения с одногнездными вместилищами диплоидные. В результате мейоза образуются гаплоидные зооиды, которые прорастают в гаплоидные растения с многогнездными вместилищами, крайне редко часть зооидов сливаются попарно и снова производят диплоидные растения с одногнездными вместилищами. Зооиды, образуемые растениями с многогнездными вместилищами, функционируют как гаметы или без слияния воспроизводят материнские растения. Иногда, например у растений Тихоокеанского побережья США, смены форм развития не происходит. В природе же и в культуре растут слоевища только с одногнездными спорангиями.
Род фелъдманния (Feldmannia) имеет слоевища высотой до 3 см, которые отличаются от эктокарпуса наличием в основании вертикальных побегов и волосков вставочных зон роста, состоящих из ряда коротких клеток, а также тем, что клетки содержат дисковидные хлоропласты. Большинство видов рода фельдманния растут на водорослях.
Род сорокарпус (Sorocarpus) включает виды, которые характеризуются многогнездными спорангиями, расположенными на ветвях пучками; одногнездные спорангии неизвестны.
Род бахелотия (Bachelotia) объединяет водоросли, внешне сильно напоминающие пилайеллу. Слоевище у них в высоту достигает 3 см. Хлоропласты в клетках вертикальных побегов удлиненные и расположены одной-двумя группами, в которых ориентированы по радиусам. В клетках ризоидов хлоропласты дисковидные. У наиболее хорошо изученного вида - бахелотии антилъской (В. antillarum) половой процесс неизвестен, в то же время у нее зооспоры из одногнездных спорангиев гораздо крупнее (15-25 X 25-35 мкм) зооспор из многогнездных спорангиев (диаметром 4-6,5 мкм).
Род стреблонема (Streblonema) - пример микроскопических эктокарповых, растущих на поверхности других водорослей и лишенных вертикальных вегетативных ветвей, а если они все-таки есть, то короткие и не отличаются от стелющихся нитей. Имеются многоклеточные волоски с базальной зоной роста (рис. 121, 2).
Род энтонема (Entonema) включает эктокарповые, растущие в слоевищах других водорослей (рис. 123). Их нити растут между клетками хозяина. Органы размножения и волоски с базальной зоной роста развиваются группами на нитях, расположенных под наружным слоем клеток или под кутикулой водоросли-хозяина; по мере роста они поднимают этот слой, разрывают его и высовываются наружу. Одногнездные и многогнездные вместилища образуются на разных слоевищах водоросли-хозяина.
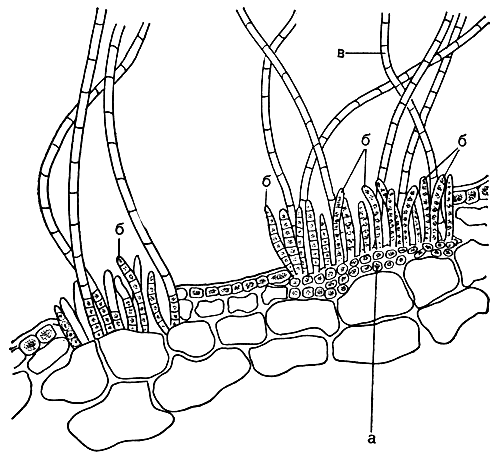
Рис. 123. Энтонема эцидиевидная (Entonema aecidioides) на поперечном срезе через пластину ламинарии: а - вегетативные клетки энтонемы; б - однорядные многогнездные спорангии; в - волоски с базальной зоной роста
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'