Общая часть
Краткая характеристика лиственных мхов
Жизненный цикл мохообразных (Bryophyta), к которому относится класс лиственных мхов (Bryopsida), характеризуется четко выраженным доминированием половой генерации - гаметофита над бесполой - спорофитом.
Развитие гаметофита начинается из споры и состоит из двух фаз развития: протонемы и гаметофора. Из прорастающей споры развивается зеленая, сильно ветвящаяся нить - протонема, представляющая собой юношескую (ювенильную) фазу развития гаметофита. Из почек, образовавшихся на протонеме, развиваются гаметофоры - листостебельные побеги, несущие половые органы. Так как на протонеме закладывается много почек, то из одной споры вырастает много гаметофоров, образующих более или менее* плотную дерновинку.
* (Дальше - б. м.)
В половых органах (гаметангиях), женских (архегониях) и мужских (антеридиях), образуются половые клетки (гаметы) - яйцеклетка и сперматозоиды. Из оплодотворенной подвижным сперматозоидом яйцеклетки развивается бесполое поколение - спорофит (спорогон), основной функцией которого является образование спор - клеток бесполого размножения.
Вегетативное тело гаметофора расчленено на стебель и листья. Корней мохообразные не имеют, а воду и растворенные в ней минеральные соли впитывают всей поверхностью вегетативного тела. Ризоиды мхов выполняют в основном функцию прикрепления гаметофора к субстрату, а в водоснабжении играют незначительную роль.
Спорофит мохообразных морфологически сильно редуцирован. Он представляет собой единичный спорангий - коробочку, сидящую на верхушке б.м. длинной, неразветвленной ножки. Ножка внизу заканчивается стопой, срастающейся с тканями верхушки гаметофора. В своем существовании спорофит связан с гаметофитом, на котором живет, за счет и с помощью которого питается.
Из спорогенной ткани в коробочке образуются споры. Споры дают начало новым гаметофитам, и цикл развития повторяется.
Споры лиственных мхов одноклеточные, преимущественно шаровидной формы. Размеры спор у представителей нашей флоры колеблются от 7 - 10 μ (у Polytrichaceae) до 200 μ (у Archidium).
Спора на поверхности покрыта 2-слойной оболочкой. Внешний слой оболочки (экзоспорий, или экзина) кутинизирован, окрашен, часто от местных утолщений скульптурный; внутренний (эндоспорий, или интина) бесцветный, целлюлозный. Споры всегда содержат протоплазму, ядро и хлоропласты, капли масла и иногда крахмал.
В благоприятных условиях освещения, влажности и реакции субстрата споры прорастают. При этом спора увеличивается в объеме, экзоспорий ее лопается, а набухшее клеточное содержимое, одетое эндоспорием, выпячивается в ростковую трубочку (рис. 1), которая вскоре отделяется поперечной перегородкой от споры и образует первую клетку протонемы. Новые поперечные деления приводят к образованию многоклеточной основной нити протонемы, обладающей неограниченным верхушечным ростом. Клетки нити могут образовывать боковые ответвления, в свою очередь многократно ветвящиеся. Вскоре после появления первой ростковой трубки на противоположной стороне споры образуется второе и даже третье выпячивание, развивающееся в новую нить, или ризоид.
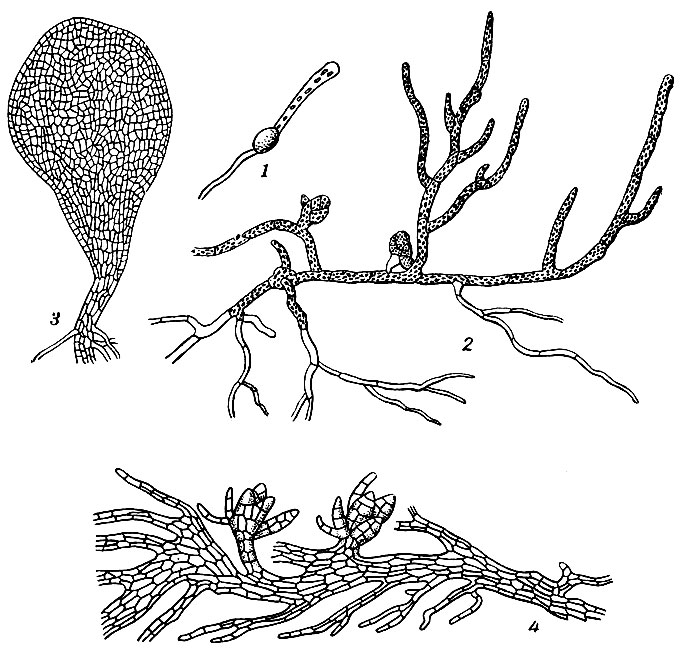
Рис. 1. 1 - прорастающая спора Funaria; 2 - нитчатая протонема Funaria с почками; 3 - пластинчатая протонема Tetraphis; 4 - пластинчатая, лопастная протонема Andreaea
Зеленые нити протонемы, стелющиеся по субстрату и пронизывающие поверхностные его слои, в совокупности образуют так называемую хлоронему, клетки которой богаты хлоропластами, имеют бесцветные клеточные оболочки и перпендикулярные к продольной оси клетки поперечные перегородки. Отходящие в глубину субстрата от протонемы многоклеточные, разветвленные и суживающиеся к концам нити - ризоиды прикрепляют протонему к субстрату и, возможно, участвуют в снабжении ее водой и растворенными в ней минеральными солями. Клетки ризоидов теряют хлоропласта, имеют только лейкопласты; их продольные стенки буровато окрашены, а поперечные перегородки расположены всегда косо к продольной оси клетки.
Наиболее распространенная форма протонемы у лиственных мхов - нитчатая; у сфагновых мхов она имеет вид пластинки; у андреевых, кроме нитей, образуются лопастные пластинки и поднятые кверху над субстратом цилиндрические стеблевидные, древовидно разветвленные образования. После некоторого периода роста на протонеме образуются почки, дающие начало листостебельным побегам. Почки закладываются на продольных стенках базальной клетки боковой ветви (реже клеток основной нити) в виде выпячивания, которое, отделившись от материнской клетки поперечной перегородкой, отчленяет одну-две клетки ножки, затем вздувается на конце и, последовательно делясь тремя косыми перегородками, образует трехгранно-пирамидальную верхушечную клетку будущего листостебельного побега. С образованием таких побегов протонема обычно отмирает. У некоторых видов она остается жизнедеятельной длительное время после образования гаметофоров или даже в течение всего цикла развития, принимая на себя функции питания не только гаметофора, но и спорофита (виды Buxbaumia).
Протонема, развивающаяся из споры, называется первичной, а образовавшаяся из частей гаметофита или спорофита - вторичной Вторичная протонема морфологически не отличается от первичной.
Листостебельный побег лиственных мхов развивается из почки на первичной или вторичной протонеме. Взрослые стебли отдельных видов мхов отличаются размерами, характером роста, способом ветвления, анатомическим строением и т. п. Рост стебля в длину осуществляется верхушечной клеткой. Длина стебля от крошечных размеров у Buxbaumia, Ephemerum, Archidium, Acaulon достигает 30 - 40 см у Polytrichum, Fontinalis и др. Вторичной меристемы у мхов нет, и стебель растет в толщину только в результате увеличения размеров меристематических клеток верхушки стебля. Диаметр стебля мхов редко превышает 1 мм.
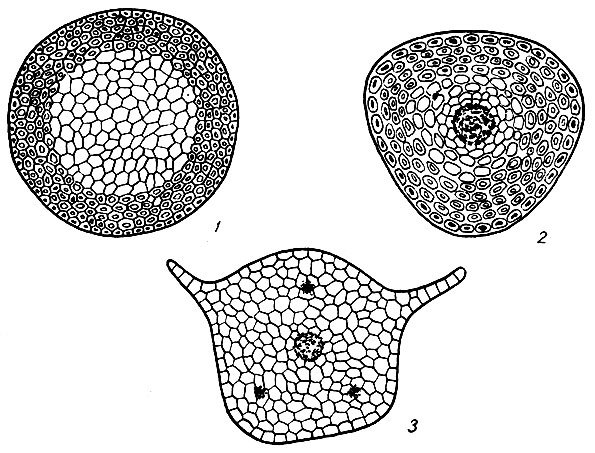
Рис. 2. Поперечные разрезы стебля: 1 - Fontinalis antipyretica, 2 - Polytrichum commune, 3 - Mnium cuspidatum
Преобладающая форма очертания стебля мхов на поперечном разрезе округлая; часто она бывает слабо-3-гранной (Plagiopus, Polytrichum), реже - овальной (Fissidens, Distichium) или округло-5-многогранной (Bartramia, Mnium) (рис. 2).
По характеру роста стебли лиственных мхов делят на ортотропные и плагиотропные. Ортотропные стебли с радиальной симметрией растут прямо вверх, архегонии и спорогоны располагаются у них на верхушке главного стебля или его равноценного ответвления. Характерны они для большинства верхоплодных (акрокарпных) мхов. Плагиотропные стебли с двусторонней симметрией плоско прижаты к субстрату, стелются по нему; архегонии и спорогоны располагаются у них на верхушке укороченных боковых ответвлений или вдоль стебля, но не на его верхушке. Свойственны они большинству бокоплодных (плеврокарпных) мхов.
Стебли лиственных мхов ветвятся. Ветвление бывает двух типов: симподиальное и моноподиальное. Симподиальный тип ветвления распространен у акрокарпных мхов, у которых рост главной оси побега прекращается с образованием гаметангиев на его верхушке. Развивающиеся непосредственно под гаметангиями новые побеги, так называемые подверхушечные (субфлоральные), или иннвации, растут в направлении главной оси, обычно не отличаются развитием от основного стебля и на верхушке заканчиваются новыми гаметангиями. Если под каждым гаметангием в нескольких последующих генерациях вырастает только один подверхушечный побег, то с течением времени образуется как бы один стебель, состоящий из субфлор ал ьных побегов, последовательно в нарастающем порядке сменяющих друг друга. Субфлоральных побегов иногда бывает больше одного, при двух - образуется вильчатое, при нескольких - пучковидное или мутовчатое ветвление.
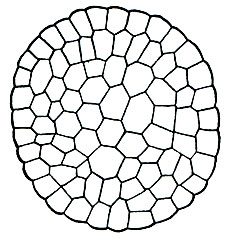
Рис. 3. Поперечный разрез стебля Andreaea
Для бокоплодных мхов характерно ветвление моноподиального типа, с четко выраженным главным стеблем и боковыми ветвями. Ветви закладываются сбоку, под растущей верхушкой стебля. Более молодые ветви всегда ближе к верхушке. Стебли бокоплодных мхов неравномерно разветвленные или при правильном чередовании ветвей - правильно перисто разветвленные. Ветви обладают ограниченным верхушечным ростом и остаются простыми или, в свою очередь, ветвятся и дают ветви второго и третьего порядка, образуя просто или дважды-трижды перисто разветвленные стебли.
Кроме симподиального и моноподиального типов ветвления, при которых боковые ветви развиваются из верхушечной части стебля, у некоторых видов акрокарпных мхов встречается вставочное, или интеркалярное, ветвление, при котором боковые ветви развиваются из нижней, более старой части стебля. К этому типу ветвления относятся кор-невищевидные и столоновидные стебли Polytrichum, Climacium, Thamnium и Rhodobryum.
Ветви некоторых видов мхов (Orthodicranum, Neckera, Isoihecium) с нитевидно утончающимися мелкооблиственными верхушками называются флагеллевидными ветвями.
Анатомическое строение стебля различных мхов довольно разнообразно. Наиболее просто построен стебель у андреевых мхов: на всей площади поперечного разреза он состоит из однообразных не дифференцированных на ткани клеток (рис. 3). У большинства мхов наблюдается дифференциация стебля на ткани, направленная на создание прочности стебля и проведение ЕОДЫ и растворенных в ней питательных минеральных солей.
В стебле лиственных мхов различаем кору, основную ткань и у более сложно устроенных - центральный пучок, или центральный тяж. Кора стебля обычно многослойная, реже 1-слойная, состоит из механических (стереидных), сильно вытянутых в длину, окрашенных клеток. Оболочки клеток периферической части коры обычно сильно утолщены, иногда до полного исчезновения просвета. У некоторых видов (Meesiu longiseta, Plagiopus oederi, Philinotis caespitosa, Eucladium verticillatum, Hygrohypnum ochraceum) поверхностные бесцветные, тонкостенные клетки коры с пористыми оболочками, увеличены в размерах и образуют гиалодермис. Внутрь стебля клетки коры становятся менее толстостенными и обычно постепенно переходят в клетки основной ткани. Иногда толстостенные клетки коры резко отграничены от основной ткани стебля (Meesia longiseta).
Основная ткань у одних видов заполняет всю внутреннюю часть стебля, у других окружает цилиндром центральный тяж. Клетки основной паренхимы живые, с богатым протоплазматическим содержимым, кроме протоплазмы и ядра имеют хлоропласта, крахмал и капельки масла. На поперечных разрезах они часто колленхиматически утолщены. Среди однообразных клеток основной паренхимы у некоторых видов встречаются группы мелких, тонкостенных клеток, так называемые листовые следы, являющиеся продолжением в стебле жилки листа (рис. 4). Листовые следы, соединяющиеся с центральным тяжем (Poly trichaceae, виды Splachnum), называются настоящими, а заканчивающиеся слепо в основной паренхиме (Mnium, Bryum, Tetraplodon),- ложными.
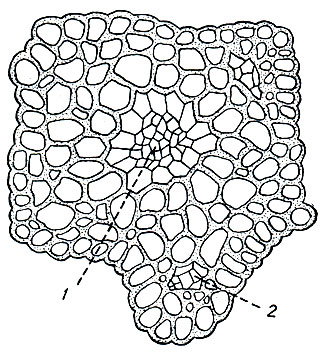
Рис. 4. Поперечный разрез стебля Mnium с простым центральным тяжем (1) и ложными листовыми следами (2)
Центральный тяж занимает осевую часть стебля. У многих видов мхов он полностью отсутствует или развит только в облиствленной части стебля (Роttiа). Клетки центрального тяжа вытянуты в длину, с косыми поперечными перегородками, преимущественно тонкостенные и бесцветные, иногда толстостенные и окрашенные (Dicranum scoparium, виды Нурпит), обычно четко отграничены от соседних клеток основной паренхимы.
Центральный тяж, построенный из однообразных клеток, называется простым, а из разнообразных, как в подземной части стебляPolytrichum,- сложным.
Стебель Polytrichum в корневищеобразной, подземной части имеет сложное анатомическое строение. На его поперечном разрезе (рис. 5) видны кора, центральный тяж и три гиподермально-радиальных тяжа, пронизывающие кору в трех местах. Периферический слой коры (эпидермис) состоит из толстостенных клеток, отдельные из которых вырастают в ризоиды. Под ним залегают два слоя паренхиматических, тонкостенных клеток и один слой крупных, тонкостенных, радиально вытянутых клеток (эндодерма). Глубже расположен трехлопастный центральный тяж, толщина которого по диаметру несколько превышает толщину коровой части. Построен он из живых толстостенных, прозенхиматических клеток, среди которых группами по 3 - 4 разбросаны тонкостенные мертвые клетки - гидроиды.
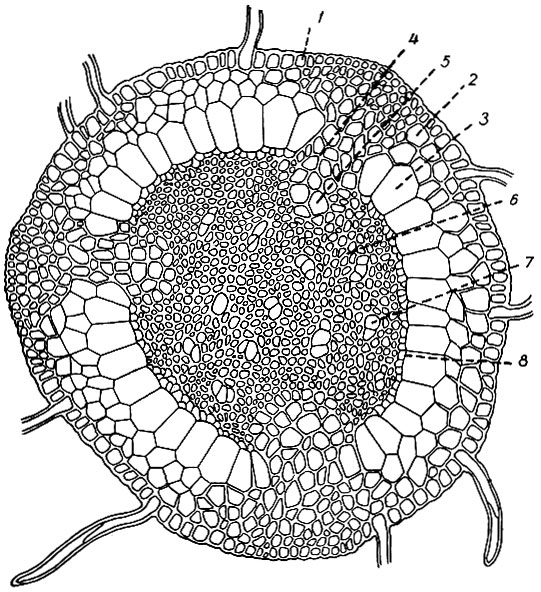
Рис. 5. Поперечный разрез подземной части стебля Polytrichum: 1 - эпидермис, 2 - паренхима, 3 - эндодерма, 4 - гиподермальный тяж, 5 - лептоида, 6 - центральный тяж, 7 - гидроида, 8 - перицикл
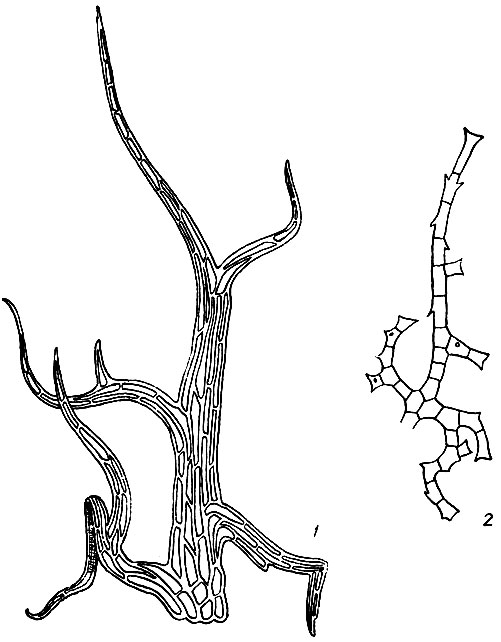
Рис. 6. Парафиллии: 1 - у Hylocomium splendes, 2 - у Thuidium
В трех местах кольцо коры прерывается гиподермально-радиальными тяжами. Гиподермальные тяжи имеют форму треугольников, обращенных основанием к периферии, а верхушками соединенных с радиальными тяжами, выходящими из углублений между лопастями центрального тяжа. Гиподермальные тяжи состоят из живых толстостенных прозенхиматических клеток, постепенно переходящих в тонкостенные паренхиматические клетки радиальных тяжей. Крайние внутренние клетки радиальных тяжей немного увеличены; обильным протоплазматическим содержимым и сквозными порами в поперечных перегородках напоминают ситовидные трубки и называются лептоидами. Центральный тяж отграничен от эндодермы тонкостенными живыми, бедными крахмалом клетками - перициклом; от радиальных тяжей, входящих в его углубления,- слоем многоугольных, богатых крахмалом клеток - амиломом. Кверху от подземной части строение стебля постепенно изменяется. Гиподермальные тяжи расширяются и сливаются в 2 - 3-слойное кольцо коры надземной части стебля. Эндодерма теряет свой крупноклеточный характер. Изменяется также строение центрального тяжа: рассеянные гидроиды соединяются в середине стебля в центральный тяж надземной части. Из богатых крахмалом клеток амилома вокруг центрального тяжа образуется гадромное влагалище, к которому снаружи прилегает лептомное кольцо, развившееся из лептоидов радиальных тяжей. За ним лежит основная паренхима с разбросанными в ней листовыми следами.
Из поверхностных клеток стебля мхов часто возникают разного рода выросты, не обнаруживающие закономерного распределения - трихомы (булавовидные волоски, ризоиды и парафилии).
Булавовидные волоски образуются в пазухах верхушечных листьев. Это - короткие, однорядные нити с булавовидно раздутой или эллиптической верхушечной клеткой.
Ризоиды представляют собой однорядные многоклеточные нити с окрашенными в коричневый, фиолетовый или красноватый цвет продольными оболочками с косыми поперечными перегородками. Образуются они из поверхностных клеток стебля у верхоплодных мхов преимущественно нижней, у бокоплодных - обращенной к субстрату части стебля. У некоторых видов ризоиды образуются из клеток жилки (Tomenthypnum) или отдельных, так называемых инициальных, клеток верхушечной части листовой пластинки (Calllergon, Drepanocladus).
Парафиллии - это зеленые, нитевидные или листовидные, простые или разветвленные выросты стебля бокоплодных мхов (Thuidiaceae, Leskeaceae, Amblystegiaceae, Rhytidiaceae и Hylocomiaceae) (рис. 6). Нитевидные парафиллии прикрепляются к стеблю одной, листовидные-двумя или большим количеством клеток своего основания.
Псевдопарафиллии возникают на стебле единично, вблизи боковых ответвлений и представляют собой недоразвитые листья.
Листья мхов всегда сидячие, бесчерешковые, обычно поперечно прикрепленные к стеблю, иногда основание листа ориентировано косо (Plagiotheciaсеае) или, как на стерильных побегах Schistostega, вдоль стебля. Листорасположение всегда спиральное, у немногих по формуле 1/2 (Fissidens, Schistostega, Distichium), чаще - 1/3 (Fontinalis, Meesia triquetra), обычно 2/5,3/8 и др.
По положению на стебле различаем: низовые, стеблевые и покровные листья.
Низовые листья размещены в подземной или нижней надземной части стебля; они обычно значительно меньше стеблевых, часто без хлорофилла.
Покровные листья, наоборот, обычно крупнее стеблевых, отличаются от них часто формой и окраской, окружают и защищают гаметангии. Листья, окружающие женские гаметангии, называются перихециальны-м и, мужские - перигониальными.
Стеблевые листья у плеврокарпных мхов часто отличаются от веточных, а у плоско облиствленных брюшные и спинные - от боковых. У верхоплодных листья б. м. одинаковые, хотя нижние обычно слабее развиты.
Листья мхов, как правило, простые, цельные, не разделены на лопасти. Сторона листа, обращенная к стеблю, называется внутренней, верхней, или брюшной, противоположная - нижней, внешней, или спинной.
В листе различаем пластинку и жилку листа. Жилка иногда слабо выражена, едва заметна или полностью отсутствует.
У преобладающего большинства мхов листовая пластинка 1-слойная; у многих видов верхняя часть (Grimmia, Syntrichia), у некоторых нижняя (Entodon), иногда край (Cinclidotus, Schistidium brunescens) и,редко вся пластинка (Grimmia unicolor) бывает дву-многослойная, причем все клетки пластинки одинаковы, равноценны. Многослойная пластинка (Leucobryum glaucum) построена из неодинаковых клеток: больших, бесцветных, тонкостенных, пористых - во внешних слоях и мелких, угловатых, хлорофилло-носных, размещенных в один слой в центре. Иногда единичные клетки или группа клеток 1-слойной пластинки делятся в плоскости пластинки и становятся дву -многослойными. В таких местах на пластинке образуются бугорки, валики или зубчики (на спинной стороне листа Atrichum).
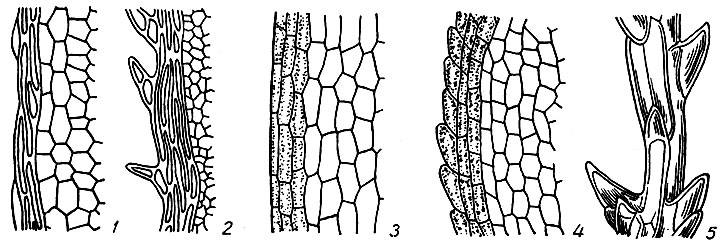
Рис. 7. Край листа: 1 - цельный окаймленный, 2 - зубчатый, 3 - цельный, 4 - городчатый, 5 - парнозубчатый
В листовой пластинке различаем основание, верхушку, край, кайму и листовые крылья, или листовые ушки. Основанием листа называется место прикрепления его к стеблю. Оно может быть суженным или расширенным, закругленным или сердцевидным, иногда низбегающим по стеблю или влагалищным. Противоположный основанию конец листа называется верхушкой, которая бывает острой, заостренной, иногда волосовидной, тупой, закругленной, б. м. выемчатой, клобуковидной, прямой, серповидно изогнутой или отогнутой назад. Края листа (рис. 7) цельные или гладкие без каких-либо выступов наружу, городчатые - с тупыми выступами верхних углов краевых клеток, зубчатые или пильчатые - с б. м. острыми, одно -многоклеточными зубцами, плоские, загнутые или завороченные внутрь, отогнутые или отвороченные назад. Кайма листа образуется одним либо несколькими рядами его краевых клеток, отличающихся формой или окраской от остальных клеток пластинки. Клетки каймы преимущественно узкие и длинные, тонко- или толстостенные, бесцветные или окрашенные, часто желтоватые или красноватые. Кайма одно- или многослойная, плоская и вздутая. Последняя может быть образована клетками, не отличающимися ни формой, ни окраской от остальных клеток листовой пластинки (Cinclidotus). Листовыми ушками называем обычно б. м. многочисленные группы клеток в углах основания листа, отличающихся формой, окраской, размерами и утолщениями клеточных оболочек от соседних клеток пластинки.
По форме пластинки листья бывают округлые с очертанием, близким к кругу; яйцевидные, когда пластинка у основания шире, чем на верхушке, а длина ее в 2 раза превышает ширину; э ллиптические - к основанию и верхушке суженные, при соотношении длины к ширине 1 : 2; ланцетные - от расширенного основания постепенно суженные к верхушке, а длина в 3 - 4 раза превышает ширину; языковидные - с закругленной или тупо заостренной верхушкой; шпателевидные - расширенные кверху; линейные - почти до верхушки равноширокие; шиловидные - с длинной, острой, иногда нитевидно вытянутой верхушкой.
Поверхность листа плоская или вогнутая, желобчатая или килеватая, продольноскладчатая или поперечноволнистая, морщинистая.
Форма листа, толщина его пластинки, характер края (зубчатый, гладкий, окаймленный, плоский, отвороченный) имеют важное диагностическое значение.
Внешний облик, расположение и ориентировка листьев в сухом и влажном состоянии различны и при определении мхов часто являются важным признаком.
В сухом состоянии листья извилистые, кудрявые, прижаты к стеблю, загнуты внутрь, спирально закручены вокруг стебля, оттопыренные. Во влажном состоянии они прижаты к стеблю, черепитчатые или отклоняются от него под определенным углом. Если листья с осью стебля образуют небольшой угол, они называются прямо отстоящими или отклоненными; если угол ближе к 45° - далеко отклоненными, если угол от 45 до 90°-оттопыренными, и при тупом угле - оттопыренно отогнутыми или отогнутыми.
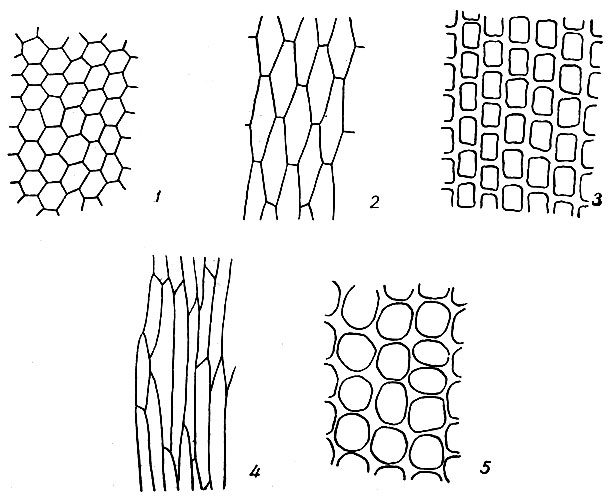
Рис. 8. Клетки листа: 1 - 3 - паренхиматические, 4 - прозенхиматические, 5 - колленхиматические
Форма клеток, слагающих пластинку листа, имеет большое значение при определении мхов. Различаем два типа клеток листовой пластинки: паренхиматические и прозенхиматические (рис. 8). Паренхиматические клетки почти одинаковые в длину и ширину: округлые, квадратные, шести - многоугольные или при значительном удлинении прямоугольные до линейно-прямоугольных, на обоих концах поперечно притупленные. Прозенхиматические клетки вытянуты в одном направлении, узкие, с заостренными концами (ромбические, линейные или червеобразные). Клеточные оболочки тонкие, равномерно или же неравномерно утолщенные. Клетки с 3-угольными утолщениями по углам называются коллен-химатическими. Иногда продольные стенки более утолщены, чем поперечные, и наоборот. У Rhacomitrium и некоторых видов Grimmia продольные стенки неравномерно выемчато утолщенные. Иногда отдельные, небольшие участки внутренних стенок клеток остаются не утолщенными. Такие стенки называются пористыми (некоторые виды Dicranum, Hypnum).
Листовая пластинка гладкая или от неровностей внешних стенок клеток б. м. шероховатая с обеих сторон или только с одной. Неровности бывают двух видов: папиллы, представляющие собой бородавчатые утолщения наружных клеточных стенок, и мамиллы - конические выступы клеток без утолщений клеточной оболочки (рис. 9). Папиллы образуются только на клетках верхней и средней частей листа. Папиллозность и мамиллозность пластинки листа имеют важное диагностическое значение.
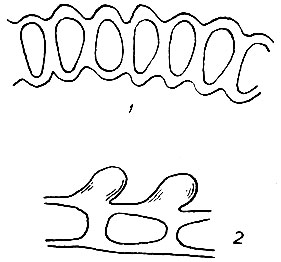
Рис. 9. Мамиллы (7) и папиллы (2)
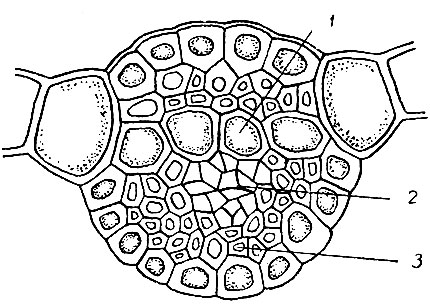
Рис. 10. Поперечный разрез жилки: 1 - указатели, 2 - спутники; 3 - стереиды
Жилка листа простая, от основания или вверху вильчато раздвоенная или едва намеченная, двойная. Простая жилка заканчивается на разном расстоянии до верхушки листа, в верхушке листа или выступает из нее в виде острия или длинного, гладкого или зубчатого, окрашенного либо бесцветного волоска. У некоторых видов верхоплодных мхов (Andreaea, Leucobryum, Ephemerum, Tetrodontium) и большего числа бокоплодных мхов жилки нет.
Жилка листа всегда многослойная. У большинства бокоплодных мхов она слабо развита, гомогенна - состоит из одинаковых клеток. У верхоплодных мхов жилка обычно хорошо развита и имеет сложное анатомическое строение (рис. 10). На поперечном разрезе жилки верхоплодных мхов хорошо заметен один, реже два ряда больших, тонкостенных, бесцветных, бедных плазматическим содержимым клеток, пронизывающих жилку поперек от основания одной половины пластинки к основанию другой и являющихся как бы продолжением клеток пластинки в жилке. Своей формой и порами на продольных стенках они напоминают гидроиды центрального тяжа. Эти клетки называются указателями и проходят либо посредине жилки (срединные), либо лежат на верхней ее стороне (брюшные). Снизу к указателям прилегает группа мелких, очень тонкостенных, богатых прото плазматическим содержимым клеток, называемых сопроводителями, или спутниками. Своим характером они напоминают лептоиды и встречаются только в жилке тех видов, которые имеют центральный тяж.
Указатели и спутники называются характерными клетками жилки. К ним обычно со спинной стороны или с обеих сторон, реже только с брюшной, прилегают клетки с сильно утолщенными стенками - с тереиды. Внешние клетки жилки, лежащие на спинной или брюшной ее стороне, имеют обычно более широкий просвет, чем стереиды, и развиты только с одной стороны жилки.
У представителей семейства Polytrichaceae и некоторых видов Pottiaсеае на верхней стороне жилки находятся зеленые выросты в виде 1-слойных, б. м. высоких, продольных пластинок (Polytrichaceae), иногда имеющих листовидный облик (Pterygoneurum) или разветвленных нитей (Aloina, Crossidium). Пластинчатые выросты образуются иногда не только на брюшной, но и на спинной стороне жилки и пластинки (Oligotrichum) или только на спинной стороне жилки (Dicranum scoparium, D. rugosum).
Наличие или отсутствие жилки листа, степень и характер ее развития являются важными систематическими признаками.
Мужские половые органы мохообразных называются антеридиями, женские - архегониями. Половые органы обычно собраны группами по нескольку в согаметангии, окруженные специализированными покровными листьями. В согаметангиях, кроме половых органов, часто находятся нитевидные, членистые образования - парафизы. Согаметангии бывают раздельнополые, состоящие только из антеридиев или только из архегониев, и обоеполые - из антеридиев и архегониев, смешанныхдруг с другом в общей обвертке (рис. 11). Согаметангии, в которых имеются архегоний, называются перихециями; мужские согаметангии, состоящие только из одних антеридиев,- перигониями.
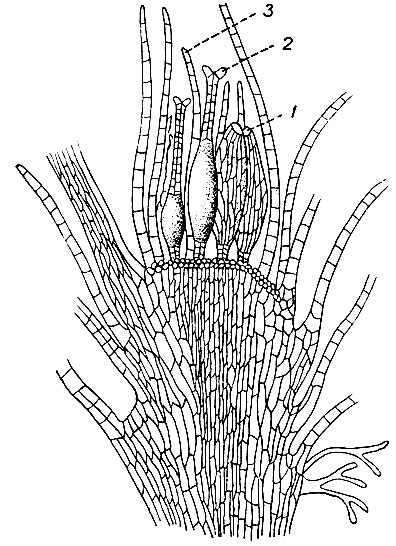
Рис. 11. Обоеполый согаметангии Bryum bimum: 1 - антеридий, 2 - архегоний, 3 - парафиза
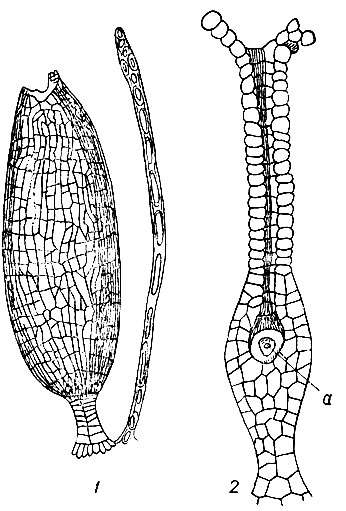
Рис. 12. Половые органы: 1 - антеридий, 2 - архегоний с яйцеклеткой (а)
Антеридий - мужской половой орган, имеет форму яйцевидного или булавовидного пузырька с 1-слойной стенкой, сидящего на короткой ножке (рис. 12). В материнских клетках сперматозоидов, которыми наполнен антеридий, образуется по одному спирально завитому, двуреснитчатому сперматозоиду. Зрелый антеридий открывается на верхушке отверстием, закрытым до того крышечкой, состоящей из изодиаметрических клеток, с сильно утолщенными стенками. Через отверстие на верхушке антеридия сперматозоиды выходят наружу и, передвигаясь в воде, устремляются к яйцеклетке в архегоний.
Архегоний - женский половой орган, бутылковидной формы, сидит на б. м. массивной ножке (см. рис. 12). Он состоит из нижней расширенной части - брюшка и верхней суженной, цилиндрической шейки. Стенка брюшка 2-слойная, шейки - 1-слойная. В полости брюшка лежит большая яйцеклетка, которая после оплодотворения сперматозоидом развивается в зародыш и дает начало спорофиту. Выше яйцеклетки лежит большая брюшная канальцевая клетка, за ней вверх по оси шейки размещены в один ряд канальцевые клетки. При созревании яйцеклетки они ослизняются. Под давлением слизи крышечка на верхушке канала открывается, слизь выходит наружу и канал шейки открывается для доступа в него сперматозоидов. В зависимости от расположения перихециев на растении мха различают две большие группы мхов: верхоплодные, или акрокарпные, у которых перихеции размещены на верхушке основного побега или его равноценного ответвления, и бокоплодные, или плеврокарпные, с перихециями на верхушке боковых укороченных ответвлений.
У некоторых видов Fissidens перихеции и спорогоны образуются посредине стебля или в его основании. У представителей родов Molendoa, Anoectangium и Cinclidotus с явно акрокарпным обликом перихеции образуются на верхушке боковых ответвлений. Виды эти образуют промежуточную группу так называемых кладокарпных видов.
По распределению половых органов на растении мха различают: одно-, дву- и многодомные виды. У однодомных видов архегонии и антеридии находятся на одном растении в раздельно- или обоеполых гаме-тангиях. У двудомных видов мужские и женские гаметангии находятся на разных экземплярах, растущих в одной или в разных дерновинках. У многодомных или одно-двудомных видов архегонии и антеридии находятся на одном растении в раздельных или обоеполых согаметангиях или же на разных растениях.
Из оплодотворенной сперматозоидом яйцеклетки развивается спорофит (спорогон). После ряда делений зиготы вычленяется двусторонняя верхушечная клетка, обеспечивающая рост спорогона в длину. Нарастая в длину, спорофит разрывает стенку архегония кольцеобразно поперек. Нижняя часть архегония, остающаяся на месте, окружает основание ножки спорогона, образуя влагалище. На верхушке влагалища у некоторых видов рода Orthotrichum развивается пленчатый, бесцветный вырост, называемый раструбом, окружающий б. м. высокой трубкой ножку спорогона. Верхняя часть брюшка архегония вместе с шейкой выносится спорогоном вверх в виде колпачка, защищающего верхушечную точку роста спорогона, а позже и молодую коробочку от неблагоприятных внешних влияний. Колпачки бывают двух типов: 1) 1-сторонние, или клобуковидные, косо сидящие на коробочке, б. м. глубоко расщепленные с одной стороны (Dicranaceae, Bryaceae, Pottiaceae, Hyp-naceae); 2) правильные, или прямостоячие,- ша-почковидные, покрывающие только верхнюю часть коробочки, и колокольчатые, покрывающие всю или почти всю коробочку, с цельным или равномерно расщепленным в нескольких местах, иногда бахромчатым нижним краем (рис. 13). Поверхность колпачка может быть гладкой или продольносклад-чатой, голой или покрытой разного рода волосками (Orthotrichaceae), образующими иногда густой войлок (Poly iridium).
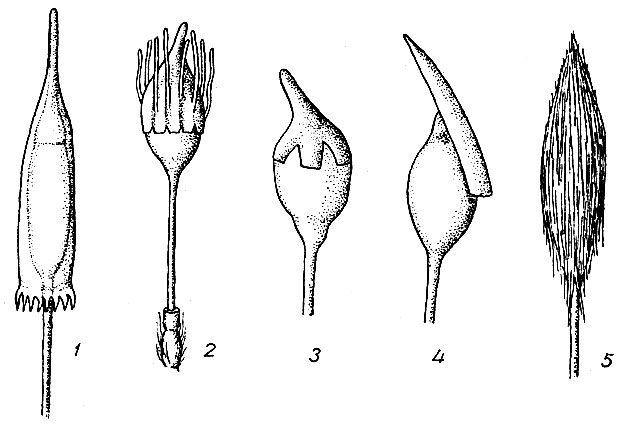
Рис. 13. Колпачки: 1 - колокольчатый у Encalypta ciliata, 2 - колокольчатый и волосистый у Orthotrichum, 3 - шапочковидный, 4 - клобуковидный, 5 - войлочный у Polytrichum
В сформированном спорогоне различают стопу, ножку и коробочку (рис. 14).
Стопа - нижняя часть спорогона, погруженная в ткани верхушки гаметофора. После созревания спорогона она вянет, сокращается, у некоторых видов спорогона легко выпадает из влагалища.
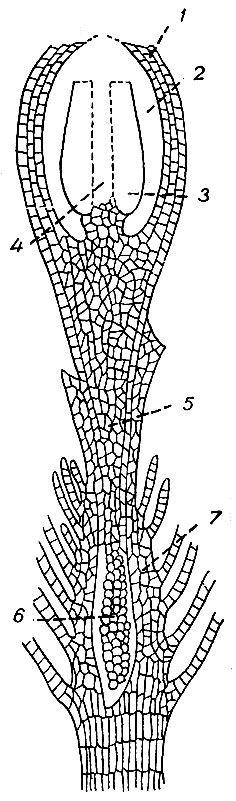
Рис. 14. Продольный разрез коробочки Orthotrichum: 1 - стенка коробочки, 2 - полость, 3 - споровый мешок, 4 - колонка, 5 - ножка спорогона, 6 - стопа, 7 - влагалище
Ножка спорогона, несущая на верхушке коробочку, цилиндрической формы, всегда нераз-ветвленная, простая. Закладываясь у всех видов, у некоторых она не развивается, и коробочка или сидит непосредственно на влагалище (Archidium), или поднимается над перихециальными листьями на удлиненной, безлистой верхушке стебля, так называемом псевдоподии (Andreаеа). При незначительной длине ножки коробочка спрятана в перихециальных листьях, при достаточной - возвышается над ними. Ножка спорогона прямая или согнутая, извилистая, часто спирально скрученная. На поперечном разрезе ножки осевую часть занимает центральный цилиндр, состоящий из тонкостенных, вытянутых по оси ножки клеток. Его окружает основная паренхима, составляющая основную массу тела ножки. На периферии ножка покрыта одним или несколькими слоями прозенхиматических стереидных клеток, окрашивающихся при созревании спорогона в красный или желтый цвет. Поверхность ножки гладкая или шероховатая от простых или разветвленных борода-вочек.
Коробочка - главная часть спорофита. Она может быть прямой - с радиальной симметрией, согнутой обычно на спинке сильнее - с двусторонней симметрией и дор-совентральной, когда верхняя и нижняя части иначе построены. По форме коробочка шаровидная, овальная, яйцевидная, цилиндрическая, грушевидная, булавовидная, призматическая и т. д. В коробочке различают шейку, гипофизу, урну, колечко и перистом.
Шейка - это нижняя массивная часть коробочки, б. м. постепенно переходящая в ножку. Она или очень длинная, превышающая длину урны, или короткая, незаметная, гладкая, продольноскладчатая или морщинистая, иногда иначе окрашенная, чем остальная часть коробочки. Иногда шейка с одной стороны вздутая и образует зобик (некоторые виды Dicranella, Oncophorus и Kiaeria).
Гипофиза, или апофиза, представляет собой б. м. значительное вздутие нижней части коробочки или верхней части ножки спорогона, резко отграниченное от урны. У Polytrichum гипофиза имеет вид кольцевидного вздутия. В семействе Splachnaceae гипофиза шире урны, зонтиковидная, бутылковидная или шаровидная, иногда ярко окрашенная.
Урной называется средняя часть коробочки (между шейкой и крышечкой), в которой образуются и сохраняются споры. Верхний край урны называется устьем; он часто несет зубцевидные выросты, образующие в совокупности перистом.
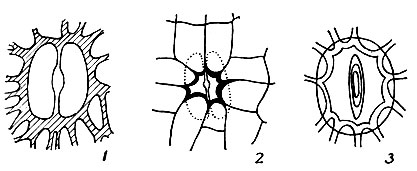
Рис. 15. Устьица: 1 - одноклеточное, 2 - погруженное, 3 - поверхностное.
На поперечном разрезе урны хорошо заметны многослойная стенка и внутренняя часть, разделенные воздушной полостью. Форма клеток эпидермиса различная и часто служит диагностическим признаком при определении. Иногда в эпидермисе коробочки продольные полоски тонкостенных клеток чередуются с полосками толстостенных, выступающих в сухих коробочках в виде продольных жилок (Orthotrichaceae). В эпидермисе коробочки размещены устьица (рис. 15). Сосредоточены они в основном в нижней части коробочки - на шейкеurипофизе, иногда рассеяны по всей стенке коробочки, Устьица бывают преимущественно дву-, реже одноклеточные (Funaria). Устьица, размещенные на одном уровне с соседними клетками эпидермиса, называются поверхностными; лежащие ниже уровня соседних клеток - погруженными.
Непосредственно под эпидермисом залегают один-два слоя бесцветных тонкостенных паренхимных клеток, образующих водную ткань. Глубже лежат хлорофиллоносные клетки, граничащие с воздушной полостью. Воздушную полость пронизывают простые или разветвленные зеленые нити, соединяющиеся с зелеными клетками внешней стенки спорового мешка. Хлорофиллоносные клетки стенки коробочки и спорового мешка образуют ассимиляционную ткань типа губчатой паренхимы.
Внутреннюю часть коробочки составляет споровый мешок, наполненный спорообразующей тканью - археспорием, и колонка. Стенка спорового мешка состоит из одного, редко двух слоев тонкостенных хлорофиллоносных клеток, теряющих ко времени созревания спор живое содержимое и легко разрушающихся. Большинство Polytrichaceae имеет еще одну, внутреннюю полость между споровым мешком и колонкой.
Колонка занимает осевую часть коробочки и представляет собой цилиндрический тяж стерильной ткани, который тянется от основания урны до крышечки. В основании и на верхушке она соединяется с тканями коробочки и у всех настоящих мхов (за исключением Archidium, у которого колонки мет) пронизывает споровый мешок. У андреевых мхов колонка не доходит до крышечки и споровый мешок покрывает ее сверху в виде купола подобно тому, как у сфагновых мхов. При созревании спор у стегокарпных мхов колонка или отделяется от крышечки (Hedwigia, Schistostega), или разрывается на две части. Линия разрыва может проходить в разных местах колонки: у многих плеврокарпных видов на уровне верхушки зубцов перистома, у большинства акрокарпных на уровне нижнего края крышечки, у Schistidium, Climacium - при основании урны. Верхняя часть колонки отпадает вместе с крышечкой, нижняя обычно сильно см орщивается и остается на дне урны в виде маленького комочка. У Splachnaceae стенка коробочки см орщивается сильнее, чемколонка, которая после отпадания крышечки' выступает из урны; у Polytrichaceae колонка вверху расширяется и образует круглую бледную мембрану - эпифрагму, закрывающую отверстие урны.
Крышечка - это б. м. четко отграниченная верхняя часть коробочки, построенная из клеток, формой и расположением отличающихся от клеток соседних частей коробочки. В зависимости от того, имеет ли коробочка крышечку, которая отпадает или нет, а также от того, как освобождаются споры из коробочки, различают стего-, клейсто-и схизокарпные мхи.
У стегокарпных видов при созревании спор крышечка отпадает и споры высыпаются через отверстие урны.
Клейстокарпные виды не имеют крышечки (Archidium, Pseudophemerum, Phascum, Acaulon) или крышечка намечена, но не отпадает (Pottia bryoides, Astomum). У них споры освобождаются из коробочки после неправильных разрывов или сгнивания ее стенок. У схизокарпных мхов (Andreaeaceae) коробочка без крышечки; при созревании спор она растрескивается 4 - 6 продольными щелями на створки, соединенные у верхушки и у основания. Крышечка бывает плоской, выпуклой, тупо-конической, на верхушке с боро-давочкой или б. м. длинным клювиком.
Колечком называется поясовидная полоска из одного или нескольких рядов крупных, уплощенных, сильно гигроскопических клеток, лежащих между верхним краем урны и крышечкой (рис. 16). Клетки колечка при колебаниях влажности изменяются в размерах и отрываются от крышечки (которая при этом отпадает) обычно и от края урны. Колечко отваливается по частям или полностью, в последнем случае часто спирально скручиваясь. Иногда колечко отрывается только от крышечки и остается при урне. У многих стегокарпных видов колечка нет.
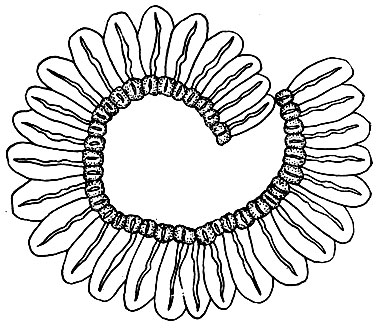
Рис. 16. Колечко
Перистомом называется совокупность зубцевидных, реснитчатых или нитевидных выростов, окружающих устье урны и составляющих своеобразный, характерный только для настоящих мхов, аппарат для регулирования высева спор. Почти все стегокарпные виды имеют перистом. Те, у которых его нет, называются гимностомными, или голоустыми. Перистом бывает простым или двойным. Простой состоит из одного ряда зубцевидных выростов, двойной - из двух концентрических рядов выростов. Внешний ряд называется наружным перистомом, или экзостомом, внутренний - внутренним перистомом, или эндостомом.
Зубцы эндостома образуются из утолщенных оболочек клеток перистом-ного слоя, залегающего непосредственно под крышечкой. Утолщения могут охватывать всю оболочку клетки или только ее часть, причем неутолщенные части оболочки разрушаются. В связи с разным характером утолщений оболочек клеток перистомного слоя лиственные мхи разделяют на две подгруппы: нечленистозубые (Anarthrodontei, или Nematodontei), у которых зубцы построены из целых, утолщенных мертвых клеток и образуются из участков ткани перистомного слоя (Polytrichaceae, Georgiaceae) (рис. 17), и членистозубые мхи (Arthrodontei), у которых зубцы состоят из утолщенных частей клеточных оболочек.
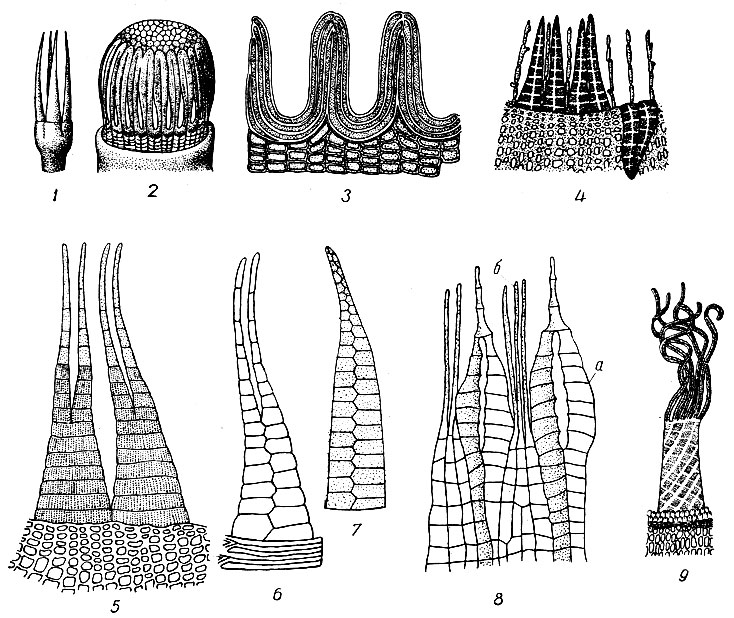
Рис. 17. Типы перистомов: 1 - Tetraphis pellucida; 2 - Polytrichum с эпифрагмой; 3 - зубцы перистома Polytrichum; 4 - Orthotiridium; 5 - зубцы перистома гаплолепидного типа с наружной стороны (Dicranum); 6 - зубец перистома гаплолепидного типа с внутренней стороны; 7 - зубец наружного перистома диплоле-пидного типа с наружной стороны {Mnium); 8 - внутренний перистом у Mnium; a - отросток, б - реснички; 9 - перистом у Syntrichia
В группе членистозубых утолщается только одна из тангентальных стенок и часто небольшие, соседние с ней участки поперечных перегородок клеток перистомного слоя. Неутолщенные участки оболочек разрушаются и от клеток остаются только отдельные, у разных видов различной формы, пластинки-членики, по-разному соединенные между собой в зубцы, постоянной для определенных видов формы.
Группа членистозубых мхов объединяет виды с простым и двойным перистомом. В первом случае перистомный слой состоит из 2, во втором - из 4 концентрических рядов клеток. Зубцы простого и наружного перистома образуются из внутренних тангентальных стенок клеток внешнего круга, образующих внешнюю поверхность зубца, и наружных тангентальных стенок внутреннего круга, образующих его внутреннюю поверхность. Охваченные утолщениями участки поперечных перегородок образуют б. м. сильно выдающиеся поперечные пластинки зубцов. Наружная поверхность зубцов в редких случаях бывает гладкой, обычно она от локальных утолщений папил-лозная, бородавчатая, продольно, косо или поперечно исчерчена. Каждый зубец, таким образом, представляет собой колонку из утолщенных частей оболочек клеток перистомного слоя, границы между которыми заметны в микроскоп в виде продольных и поперечных линий на поверхности зубца. В строении отдельного зубца принимает участие обычно три ряда клеток: два из наружного круга и один из внутреннего или, наоборот, один из наружного и два из внутреннего (см. рис. 17). Исходя из этих различий, среди членистозубых мхов выделяют: 1) гаплолепидные (Haplolepideae), у которых наружная поверхность зубца состоит из одного ряда пластинок, а внутренняя - из двух рядов, соединяющихся посредине зубца; к ним относятся порядки Fis-sidentales, Dicranales, Pottiales и Grimmiales, имеющие простой перистом; 2) диплолепидные (Diplolepideae), имеющие обычно двойной перистом, наружная поверхность зубца, экзостома которого состоит из двух рядов пластинок, а внутренняя - из одного; 3) гетеролепидные (Heterolepideae) с одним семейством Encalyptaceae, у представителей которого простой или двойной перистом, построенный по гаплолепидному или диплолепидному типу.
Простой перистом и экзостом двойного перистома состоят из постоянного, для отдельных видов характерного числа зубцов: 4, 8, 16, 32 или 64. По форме зубцы бывают ланцетные, шиловидные, нитевидные, линейные, цельные, нерасщепленные или только вверху или же до самого основания расщепленные на 2 - 3, иногда нитевидные, доли. В простом перистоме зубцы иногда бывают попарно (Orthotrichaceae) или удвоенно парно (Splachnaceae) соединены, иногда срастаются основаниями друг с другом и образуют общую основную перепонку, у отдельных видов едва выступающую над краем урны, у других значительно его превышающую, а у видов Syntrichia достигающую 2/3 высоты перистома (см. рис. 17). В двойном перистоме внутренний перистом построен сложнее наружного. Обычно он короче наружного, только у Amblyodon, Meesia и Cinclidium длиннее его. Иногда бывает слабо развитым - рудиментарным (Leucodon, виды Neckera, Anomodon) или состоит из отдельных ресничек, чередующихся с зубцами экзостома (Orthotrichaceae) (см. рис. 17), или ланцетных выростов, противостоящих зубцам экзостома (Funaria). У Fontinalis внутренний перистом образует решетчатый конус, а у Diphyscium и Buxbaumia - продольноскладчатую, коническую, пленчатую трубку. Вполне развитый внутренний перистом (Bryaceae, Mniaceae, Нурnасеае) состоит из б. м. высокой килевато-складчатой основной перепонки (мембраны), на которой между каждыми двумя зубцами экзостома стоят ланцетные отростки, обычно вдоль середины щелевидно или окновидно продырявленные. Между отростками на перепонке сидят по 2-3 реснички (см. рис. 17). Реснички гладкие или папиллозные, узловатые или с боковыми привесками. Иногда они бывают слабо развитыми - рудиментарными или отсутствуют.
У некоторых видов Grimmia, Encalypta и Orthotrichum встречается пред-перистом. В редких случаях предперистом состоит из правильных зуб-цевидных выростов, образующих третий круг; в большинстве же случаев он представлен пластинчатыми утолщениями неправильной формы, прилегающими снаружи к зубцам перистома.
Внешние стенки зубцов перистома обычно утолщены сильнее, чем внутренние. Различия в утолщениях стенок зубцов и способность их быстро набухать или см орщиваться при изменениях влажности обусловливают гигроскопические движения зубцов. При увлажнении зубцы обычно куполовидно сходятся над отверстием урны, закрывая выход спорам и препятствуя попаданию в урну воды, при подсыхании - выпрямляются, звездчато расходясь, или отгибаются, открывая устье урны.
Многие, в частности двудомные, виды лиственных мхов спорогоны образуют редко или встречаются только в стерильном состоянии. Распространение и сохранение этих видов в пределах занятой ими территории возможно только благодаря их способности к вегетативному размножению.
Известно, что почти каждая клетка вегетативного тела лиственных мхов может образовывать протонему и на ней новые растения. Кроме того, у лиственных мхов часто встречаются специальные выводковые органы, образующиеся на стеблях, листьях, протонеме или ризоидах (рис. 18). Известны следующие выводковые органы: ломкие ветви, выводковые ветви, выводковые почки, выводковые клубеньки, ломкие листья, выводковые листья и выводковые тельца.

Рис. 18. Выводковые органы: 1 - выводковая почка Bryum argenteum; 2 - побег В. argenteum с выводковыми почками; 3 - выводковая почка Pohlia bulbifera; 4 - выводковый клубенек Leptobryum pyriforme; 5 - выводковые тельца Syntrichia papulosa; 6 - выводковое тельце Bryum erythrocarpum; 7 - верхушка листа Leucobryum glaucum; 8 - выводковые тельца Tayloria serrata; 9 - выводковые тельца Zygodon vlrldlssimus; 10 - выводковые нити Encalypta streptocarpa; 11 - выводковая веточка Hopteryglum elegans; 12 - корзинка с выводковыми тельцами Tetraphls pelluctda; 13 - продольный разрез через корзинку Т. pelluclda
Ломкие ветви, или флагеллы,- нитевидные, с мелкими, чешуйчатыми листьями побеги, собранные по несколько в верхней части стебля (Orthodicranum flagellare, Isopterygium elegans), в зрелом состоянии они легко отламываются от материнского растения, распадаются на части, каждая из которых способна образовывать протонему. Выводковые ветви отличаются от ломких ветвей тем,что,отделившись от материнского растения, не распадаются на части, а остаются целыми, образуют ризоиды и развиваются в нормальные особи (Neckera complanta, N. pumila, Leucodon sciuroides, Grimmia incurva, Mniobryum wahlenbergii). Выводковые почки близки к выводковым ветвям и представляют собой сильно укороченные листостебельные побеги (Anomobryum, Bryum argenteum, Leskeella), иногда с мелкими (некоторые виды Pohlia), едва намеченными (Pohlia proligera) листочками. Образуются они единично или скученно в пазухах листьев. Выводковые клубеньки образуются при дальнейшем укорачивании продольной оси, сопровождающемся увеличением ее объема и полной редукцией листьев (Leptobryum pyriforme). Ломкие листья отличаются от нормальных тем, что их верхушки легко отламываются и дают протонему (Dicranum fragilifolium, D. viride, Orthodicranum striatum, Tortella tortuosa). Выводковые листья, внешне не отличающиеся от обычных, отпадают от стебля при легком прикосновении, в то время как обычные отделяются с трудом. Образуются они на стебле периодически, отчего стебель выглядит как бы прерывисто облиственным, как у Dicranodontium denudatum. Выводковые тельца обычно представляют собой многоклеточные образования различной формы: шаровидные (Bryum erythrocarpum, Didymodon rigidulus, Grimmia anomala, G. hartmanii), нитевидные (Orthotrichum lyellii, Amblystegium compactum), цилиндрически булавовидные (Gyroweisia, Aulacomnium), эллиптические (Orthotrichum), линзовидные (Tetraphis). Выводковые тельца образуются единично или скученно на стебле, в пазухах листьев, на листовой жилке, на пластинке листа или на ризоидах. У Tetraphis выводковые тельца собраны в корзинковидной обвертке на верхушке стебля, у Aulacomnium собраны в головку на конце безлистного побега, вырастающего на верхушке стебля.
Запчасти Daewoo на sibmashineri.ru.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://volimo.ru/ 'Водоросли, лишайники, мохообразные в природе и промышленности'